Page 1
UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR
ÁREA INTERDICIPLINARIA DE CIENCIAS DEL MAR
DEPARTAMENTO DE BIOLOGÍA MARINA
TESIS
ESTUDIO DE LA ESTRUCTURA ANTIGÉNICA DE LA LARVA L3 DE
Contracaecum multipapillatum (sensu lato)
QUE COMO REQUISITO PARA OBTENER EL TÍTULO DE
BIÓLOGO MARINO
PRESENTA:
MARCO ANTONIO SALAZAR BERMÚDEZ
DIRECTORA:
M en C. MARÍA DEL CARMEN GÓMEZ DEL PRADO ROSAS
LA PAZ, BAJA CALIFORNIA SUR, MARZO 2011
Page 2
A mis padres
María Inés y Leonardo
Page 3
Agradecimientos
Antes que a nadie, a mi Maestra, María del Carmen Gómez del Prado Rosas por su invaluable apoyo a nivel profesional y personal. Por la dedicación, paciencia y congruencia con la que realiza su labor, lo cual me motiva a seguir su ejemplo y por lo que siempre estaré en deuda. Mi primer y única “mamá académica”.
Al Dr. Felipe de Jesús Ascencio Valle, por su atención y apoyo, indispensables para que este trabajo llegara a su culminación y por sus comentarios para mejorarlo. Al Dr. Robin M. Overstreet por su amable ayuda para la identificación taxonómica
Al Dr. Carlos Armando Sánchez Ortiz por su disposición y valiosos comentarios. A todos los que intervinieron directa e indirectamente en la realización de este trabajo en la UABCS, CIBNOR, y el Centro de Investigación Biomédica de Oriente, CIBIOR.
A Ma. de Jesús “Chula” Romero Geraldo por toda su ayuda y a los que fueron mis compañeros durante mi estancia en el laboratorio de Patogénesis Microbiana del CIBNOR, al Dr. Amaury Cordero Tapia y el resto del personal del bioterio.
A las personas cuya amistad me ayudó y sigue ayudando a continuar durante mis ya muchos y antisociales años en La Paz, a Paola “Prieta”, Sr. Horacio, Paty, Isa y a todas las niñas y no tan niñas del Laboratorio de Parasitología por estar ahí y por aguantarme a mí y a mi música.
A los que, estando lejos, llevo a todos lados conmigo, Eliana, Elena, Pepe, Lalo por su amistad y su apoyo.
A toda mi familia “Salazar” y “Bermúdez” y a mis abuelos Leonor y Antonio por su cariño y apoyo.
A Roberto “primature” (R-Bear), ¿qué puedo decir que no sepas ya? Por ser un amigo y un hermano indispensable para mí, todo este tiempo; por crecer y compartir lo bueno y lo malo conmigo (sobre todo lo bueno: apoyo, buena música y jocosidad). No lo hubiera logrado sin vos.
A mi mujer Ana Luisa, por ser mi motivo y voluntad día con día, para intentar ser mejor; por permitirme ser parte de tu vida y ser la mejor parte de la mía. Gracias por todos los años de felicidad que me has dado.
A mi familia; mi hermano Leonardo, mi primer ejemplo a seguir. A mis padres Leonardo y Ma. Inés por ser mi mayor ejemplo de amor, integridad y perseverancia; porque no sería lo que soy de no ser por ustedes.
Page 5
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
I
ÍNDICE
Lista de Figuras II
Lista de Tablas III
Resumen IV
Introducción 1
Antecedentes 10
Justificación 12
Hipótesis 13
Objetivo general 13
Objetivos particulares 13
Área de estudio 14
Material y Métodos 16
Recolecta de material biológico 16
Identificación de las larvas 17
Obtención del extracto crudo de las larvas 17
Obtención de suero hiperinmune 17
Análisis electroforético de los antígenos de larvas
de Contracaecum sp. 18
Inmunoblotting de los antígenos de larvas
de Contracaecum sp. 20
Resultados 22
Identificación taxonómica de hospederos y nemátodo 22
Análisis electroforético de los antígenos de larvas 23
Obtención de suero hiperinmune
e inmunoreconocimiento de los antígenos 26
Page 6
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
II
Discusión 29
Identificación taxonómica de los nemátodos 29
Parasitismo por Contracaecum 30
Estructura antigénica e inmunogenicidad
de Contracaecum multipapillatum s. l. 36
Futuro de la línea de investigación 41
Conclusiones 42
Recomendaciones 43
Trabajos Futuros 45
Literatura Citada 46
LISTA DE FIGURAS
1 . Mugil cephalus 2
2 . Ciclo de vida probable de C. multipapillatum s. l. 7
3 . Morfología de la larva L3 de C. multipapillatum s. l. 9
4 . Bahía de La Paz 15
5 . Metodología 21
6 . Perfiles proteicos del extracto crudo
de C. multipapillatum s. l. 24
7 . Antígenos de C. multipapillatum s. l. reconocidos
por anticuerpos en suero de conejo 27
Page 7
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
III
LISTA DE TABLAS
1 . Estándar de pesos moleculares 19
2 . Prevalencia e Intensidad parasitaria de C. multipapillatum s. l. 22
3 . Pesos moleculares de los
antígenos de C. multipapillatum s. l. 25
4 . Antígenos de C. multipapillatum s. l. reconocidos
por suero de conejo 28
Page 8
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
IV
RESUMEN
El nemátodo Contracaecum multipapillatum (sensu lato) pertenece a la familia
Anisakidae, cuyas larvas son un problema importante para la industria pesquera en
diferentes países, ya que son capaces de infestar el tracto digestivo del humano tras
su ingestión accidental al consumir carne de pescado cruda o poco cocinada,
produciendo la enfermedad conocida como Anisakidosis. Los nemátodos no llegan a
hacerse adultos en el humano, pero sus antígenos pueden ser reconocidos por el
sistema inmune, provocando reacciones alérgicas con niveles elevados de
Inmunoglobulinas específicas. Debido a la alta prevalencia e intensidad parasitaria
de C. multipapillatum s. l. observada en Mugil cephalus, un pez de importancia
comercial en La Paz, Baja California Sur, se realizó el estudio de la composición e
inmunogenicidad de las proteínas somáticas de larvas en tercer estadio obtenidas de
M. cephalus. Se preparó un extracto crudo de las larvas el cual se analizó por medio
de electroforesis en geles de poliacrilamida, obteniendo un perfil de 54 proteínas con
pesos moleculares entre 1.2 y 90.9 kilodaltones (kDa). Con el extracto se inmunizó a
conejos de Nueva Zelandia para obtener suero hiperinmune con Inmunoglobulinas
anti - C. multipapillatum s. l. y se analizó con la técnica de inmunoblotting; el suero
presentó IgG’s capaces de reconocer 33 de los antígenos presentes en el suero, 19
(61%) de ellos (90.94 - 8.55 kDa) reconocidos por IgG’s producidas por la presencia
de los antígenos de C. multipapillatum s. l. y 14 reconocidos por IgG’s presentes
también en el suero del conejo control. Es necesario alertar a las autoridades y a la
población en general, para que adopten medidas precautorias, con el fin de disminuir
el posible riesgo que representa C. multipapillatum s. l. a la salud pública. A partir de
la información obtenida es posible realizar un estudio que determine si existen casos
de infección o reacciones alérgicas producidas por esta especie.
Page 9
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
1
INTRODUCCIÓN
Los peces, tanto en estado silvestre como en condiciones de laboratorio,
están sujetos a ser parasitados por organismos pertenecientes a diversos grupos
taxonómicos. Las lesiones causadas por los parásitos, sobre todo en peces
silvestres, por lo general pasan inadvertidas, a menos que el daño sea tan obvio
que demerite su calidad comercial (Roberts, 1981).
Los nemátodos frecuentemente parasitan peces, en ocasiones, como parte
de ciclos de vida complejos que incluyen varios estadios larvarios. Se estima que
cerca de 40,000 especies parasitan vertebrados; se han descrito cerca de
17,000, comprendidas en 256 familias y 2,271 géneros (Anderson, 2000).
Los peces del género Mugil (Lisas) han sido estudiados en Baja California
Sur (B.C.S.) desde 1995 en relación con su helmintofauna, registrando elevada
prevalencia y carga parasitaria por larvas L3 del nemátodo Contracaecum sp.
(Gómez del Prado et al., 2008); estos parásitos parecen acumularse durante el
tiempo de vida de los peces. En el estado de Baja California se ha observado
que la intensidad parasitaria de Contracaecum spp. y otros parásitos
combinados, aumenta respecto al tamaño de Mugil cephalus, (Valles-Ríos et al.
2000).
Las especies de “lisas” son muy abundantes y valiosas en las costas de
México. En el Golfo de California en el periodo de 2001-2005 constituyeron la
segunda mayor captura (en toneladas y ganancias promedio) de la flota ribereña.
En la Bahía de La Paz, B.C.S., es posible encontrar Mugil cephalus (Fig. 1) y M.
curema a lo largo del año. A pesar de que no se encuentran entre las especies
que aportan mayor porcentaje de captura en el área, representan un recurso local
Page 10
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
2
importante debido a que se consume a nivel popular por su bajo precio. (Pérez y
Ruiz-Luna, 1985; Plomozo-Lugo, 2011).
Figura 1. Mugil cephalus “Lisa cabezona” de la Bahía de La Paz, B. C. S., México.
Las larvas de nemátodos de la familia Anisakidae en peces, son un
problema importante para las industrias pesqueras de varios lugares de Europa y
Estados unidos de América, siendo Anisakis, Contracaecum, Hysterothylacium
y Pseudoterranova los géneros más frecuentes.
La presencia de los nemátodos anisákidos adultos está asociada a
patologías ligeras en su hospedero definitivo (aves y mamíferos piscívoros). De
acuerdo con Stoskopf, 1993 y Anderson, 2000, las larvas de Anisakis y
Contracaecum pueden causar considerable daño a los órganos viscerales y otros
tejidos de su hospedero intermediario o paraténico (generalmente peces), debido
a la conducta migratoria que tienen para su establecimiento en la serosa de
varios órganos del aparato digestivo. El género Contracaecum en particular,
puede provocar serios daños en hígado y tejidos mesentéricos de sus
hospederos, tales como inflamación extensiva, fibrosis y adhesión visceral pero
Page 11
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
3
sin impacto aparente en la condición corporal. Así ocurre en las lisas en la Bahía
de La Paz, las cuales no presentan alteraciones externas visibles a pesar de las
concentraciones relativamente altas de parásitos en el cuerpo, por lo que su
presencia no afecta significativamente su comercialización, lo que constituye un
peligro potencial de presentar una reacción a los alérgenos en los alimentos que
estuvieron en contacto con las larvas o incluso, contraer la Anisakidosis.
(Mbahinzireki, 1980; Nadler et al., 2005; Gómez del Prado et al., 2008).
La anisakidosis en humanos, es una infestación parasitaria del tracto
digestivo por nemátodos de la familia Anisakidae, resultado de la ingestión
accidental de su tercer estadio larvario (L3) en carne de pescado cruda o poco
cocinada, siendo Contracaecum, Pseudoterranova, Anisakis los géneros más
frecuentes. Se reserva el término anisakiasis a la patología producida
específicamente por el género Anisakis y ambos términos son utilizados en la
literatura (Gómez Sáenz et al., 1999; Noh et al., 2003). La anisakidosis fue
endémica de Japón, pero debido al consumo mundial de pescado crudo, puede
convertirse en una enfermedad cosmopolita, a pesar de que se conoce que las
larvas se destruyen al cocer el pescado a una temperatura de 60°C o por
congelación a -20°C (Noh et al., 2003).
Los nemátodos no llegan a hacerse adultos en el humano, pero su
presencia expone al hospedero a antígenos que pueden ser reconocidos por el
sistema inmune, estos antígenos son usualmente proteínas y glicoproteínas
localizadas en la superficie o en los productos de secreción. La inmunidad contra
las infecciones por nemátodos, al igual que muchas otras helmintiasis, es en su
mayor parte humoral (Perec y Okulewicz, 2006)
Page 12
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
4
Tras la ingestión de la larva, ésta penetra activamente la pared del
estómago o del intestino delgado, causando síntomas gastrointestinales agudos
que generalmente se confunden con apendicitis o úlcera gástrica, dichos
síntomas pueden presentarse desde una hora hasta dos semanas después de
consumir el pescado contaminado (Kim et al., 1991; Noh et al., 2003). La
anisakidosis provoca episodios que pueden variar desde una ligera urticaria hasta
anafilaxia recidivante; pueden dar origen a granulomas eosinófilos en la pared
gastrointestinal, además de reacciones alérgicas de tipo 1, con niveles elevados
de Inmunoglobulina tipo E específica (IgE), diagnosticadas generalmente por
serología (Roberts, 1981; Gómez Sáenz et al., 1999; Anderson, 2000; Lorenzo et
al., 2000; Cuellar et al., 2001; Rodero et al., 2001). En algunos pocos casos, se
han encontrado larvas L3 de Anisakis simplex y de Contracaecum sp. en alvéolos
o bronquíolos produciendo hemorragia y enfisema que se ha llegado a confundir
con asma (Matsuoka et al., 1994). Desde hace varios años, en España, ha sido
posible diagnosticar varios casos de Anisakidosis gástrica e intestinal con
endoscopía y tratándose con cirugía (Sugimachi et al., 1985; Isishukura, 1989;
Sakamari, 1990)
Se ha observado que los nemátodos del orden Ascaridida, que incluye a
los anisákidos, son capaces de infectar mamíferos, aun cuando su hospedero
definitivo no sea un organismo de sangre caliente; Contracaecum multipapillatum
parasita el proventrículo de aves piscívoras y al igual que la mayoría de los
miembros del género, no afecta naturalmente a mamíferos terrestres, aunque
varias especies de Contracaecum se presentan en mamíferos marinos (Vidal-
Martínez et al., 1994). Se ha demostrado que C. multipapillatum puede madurar
Page 13
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
5
en un mamífero terrestre de manera experimental, señalando el peligro potencial
a la salud pública. Aun cuando los juveniles de esta especie se encuentran
generalmente encapsulados en hígado, riñones y mesenterio de los peces
hospederos, también pueden encontrarse en cualquier otro lugar e incluso
permanecer dentro de ellos después de eviscerar, por lo que podrían ser
ingeridos posteriormente por el humano; si bien las larvas de Contracaecum spp.
no son ingeridas tan frecuentemente como las que se presentan típicamente en
la carne de los peces (ej. Anisakis spp.), esto ha provocado que sean menos
estudiados (Fagerholm, 1988; Vidal-Martínez et al., 1994).
Contracaecum multipapillatum es un parásito de pelícanos, cormoranes y
garzas en América, aunque los registros de juveniles indican que la especie
pudiera extenderse a Asia y África (Vidal-Martínez et al., 1994). El ciclo de vida
del género Contracaecum ocurre en el agua e involucra a invertebrados acuáticos
y peces como hospederos intermediarios o paraténicos (Anderson, 2000).
Las hembras adultas depositan los huevos en el tracto digestivo para ser
expulsados junto con sus excretas. En el agua, los huevos embrionados
contienen el primer estadio larvario, éste muda poco después, dando lugar al
segundo estadio; esta larva eclosiona en el agua y es ingerida por un
invertebrado crustáceo, generalmente copépodos que actúan como primer
hospedero intermediario, la larva migra a su hemocele en donde aumenta
ligeramente su tamaño y presenta un desarrollo parcial de los órganos.
Posteriormente los peces que se alimentan de copépodos, actúan como
segundos hospederos intermediarios o paraténicos, donde alcanzan el tercer
estadio (L3), las larvas crecen notoriamente, atraviesan las paredes del intestino
Page 14
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
6
hacia la cavidad corporal y se encapsulan en el mesenterio y diferentes órganos
(hígado, gónadas, riñón); se ha demostrado que la transferencia de las larvas de
un pez a otro es posible mediante depredación. Finalmente, el pez infectado es
ingerido por el ave hospedera, las larvas son liberadas en el tracto digestivo y se
adhieren a la pared del estómago donde se alimentan de lo ingerido por el ave. El
desarrollo posterior de las larvas y paso a la adultez probablemente sucede ya
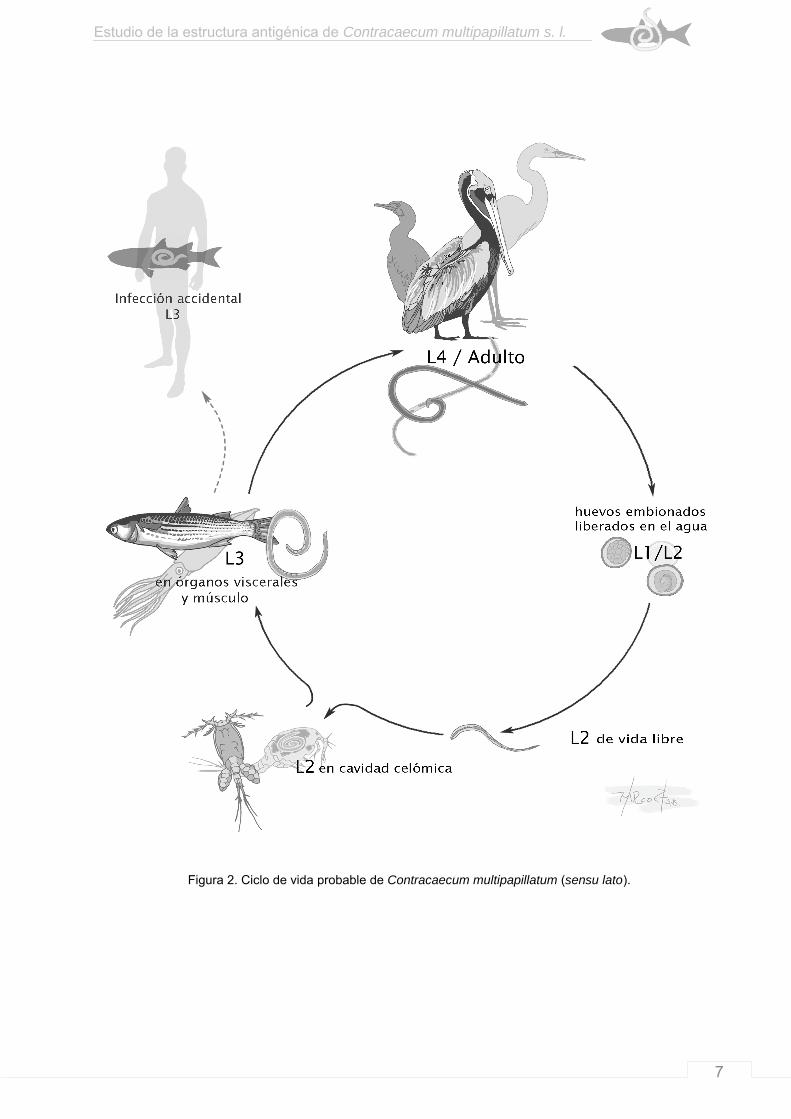
dentro del proventrículo del ave (Huizinga, 1967; Anderson, 2000) (Fig. 2).
Page 15
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
7
Figura 2. Ciclo de vida probable de Contracaecum multipapillatum (sensu lato).
Page 16
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
8
Las larvas L3 de Contracaecum miden entre 4 y 16 mm, con un extremo
posterior cónico y cola postanal con espina terminal (mucrón). Presentan una
cutícula gruesa con estriaciones transversales. En la parte anterior se encuentran
tres labios poco desarrollados (dos ventrolaterales y uno dorsal) y un diente
cuticular ligeramente romo cerca de los labios ventrolaterales y el poro excretor
inmediatamente posterior al diente. Presentan una faringe muscular y un
ventrículo glandular pequeño y esférico del que deriva un apéndice ventricular
dirigido hacia el extremo posterior. También presenta un ciego intestinal más
grande que el apéndice ventricular, dirigido hacía el extremo anterior (Berland,
1989) (Fig. 3).
El género Contracaecum Railliet y Henry, 1912 incluye aproximadamente
100 especies con una distribución mundial, la cual ocurre de manera natural
debido, probablemente, a las aves migratorias y la baja especificidad para elegir
hospederos intermediarios y definitivos (Deardorff y Overstreet, 1980; Mhaisen et
al., 1988; Anderson, 2000). Es muy complicada la identificación de anisakidos
hasta nivel especie basada en las características morfológicas de adultos
(muchas relevantes solo en machos), y es más difícil aun para las fases larvarias;
además algunos de los caracteres morfológicos que se han usado para
diferenciar especies carecen de valor interespecífico, lo que ha llevado, en parte,
a que algunas morfoespecies de éste y otros géneros de la familia Anisakidae
formen, en realidad, parte de complejos de varias especies diferenciadas
genéticamente, en ocasiones, con distribución geográfica y hospederos distintos
(Labriola y Suriano, 1996; Nadler et al., 2005; Mattiucci y Nascetti, 2008; Shamsi
et al., 2008; Shamsi et al., 2009).
Page 17
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
9
Figura 3. Morfología de la larva L3 de Contracaecum multipapillatum (sensu lato) en Mugil
cephalus. Ambos ciegos visibles (flechas). Extremo anterior con diente cuticular. Extremo
posterior con espina caudal (mucrón)
Page 18
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
10
Dentro de los complejos multiespecíficos de Contracaecum se ha
observado un nivel bajo de variación en las secuencias genéticas, por lo que es
necesario una caracterización genética que demuestre aislamiento reproductivo
entre especies que en ocasiones son simpátricas en sus hospederos definitivos
(Shamsi et al., 2009; Mattiucci et al., 2010).
Existen varios estudios de caracterización genética de las especies de
Contracaecum parásitas de aves piscívoras, en los que se utilizaron aloenzimas
y las secuencias como las del primer y segundo espaciador transcrito (ITS1,
ITS2) y las subunidades 18s y 28s del ADN ribosomal; así como el citocromo B
(Cyt-B) y citocromo oxidasa 1 y 2 (COx1, COx2) del ADN mitocondrial.
Dentro del complejo multiespecífico de C. multipapillatum propuesto por
Mattiucchi et al. (2010) se encuentran 2 especies nominales nuevas identificadas
mediante COx2: C. gibsoni y C. overstreeti en Pelecanus crispus de Grecia;
antes denominadas C. multipapillatum sp. “A” y C. multipapillatum sp. “B” por
Nascetti et al. (1990). El complejo además incluye a C. multipapillatum sp. “C” de
P. occidentalis en Colombia y C. multipapillatum sp. “D” encontrada en P.
conspicillatus en Australia y caracterizadas utilizando los ITS1 e ITS2 (Shamsi et
al., 2008; Mattiucci et al., 2006 y 2010).
ANTECEDENTES
Los casos de infección por larvas L3 de anisákidos mencionados
anteriormente, han despertado el interés por continuar investigando otros
aspectos relacionados con su sintomatología, lesiones, diagnóstico y prevención
(Kato et al., 1992; Matzuoka, 1994; Angot y Brasseur, 1995; Bouree et al., 1997;
Page 19
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
11
Noelle, 1998; Del Pozo et al., 1999; López-Penas et al., 2000; Audicana et al.,
2002).
Moreno-Ancillo, 1997; Moreno et al., 2000 y Pérez-Pérez et al., 2000
realizaron estudios sobre reacciones alérgicas y caracterización de los alérgenos
de las larvas de anisákidos. Por su parte, Gutiérrez-Ramos et al., 2000
identificaron los anticuerpos anti-Anisakis y posteriormente, Rodero et al., 2001
lograron la purificación de antígenos de Anisakis.
Por otro lado, los estudios relacionados con aspectos inmunológicos de
Contracaecum son muy escasos, Coscia y Oreste (1998) identificaron
anticuerpos específicos para proteínas de Contracaecum en diferentes
teleósteos. En México son escasos los estudios que aborden el tema de la
Anisakidosis y no existen casos reportados de esta enfermedad por ninguno de
los géneros de la familia.
En varios estados de México se ha encontrado al género Contracaecum
parasitando gran variedad de hospederos, incluyendo diversas especies de
peces (Juárez, 1985 y 1986; Juárez-Arroyo y Salgado-Maldonado, 1989; Pérez-
Ponce de León et al., 1992; Mendoza, 1994; Flores, 1995; Meléndez y Rosas,
1995; Castillo, 1996; Mendoza et al., 1996; Pérez-Ponce de León et al., 1996;
Castillo-Sánchez et al., 1998; Salgado-Maldonado et al., 2001); en aves
(Caballero, 1935; Caballero y Peregrina,1938; Bravo, 1939; Alencaster, 1948;
Caballero, 1948; Caballero, 1951; Caballero-Deloya, 1960; Amaya, 1990;
Ramos, 1994) y mamíferos (Caballero y Peregrina, 1938; Vidal-Martínez et al.,
1994)
Page 20
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
12
En el país, las especies Mugil curema y M. cephalus han sido objeto de
estudio en diversos aspectos: su parasitismo (Salgado-Maldonado y Barquín-
Álvarez, 1978; Juárez-Arroyo y Salgado-Maldonado, 1989; Valles-Ríos et al.,
2000), abundancia y pesquerías (Stuardo y Martínez, 1975; Warbuton, 1979;
Rivas, 1985; Rodríguez-Romero et al., 1994; Plomozo-Lugo, 2010), crecimiento y
sobrevivencia (Muhlia-Melo y Muhlia-Almazán, 1995) y hábitos alimenticios
(Sánchez-Rueda, 2002).
Debido a la amplia distribución que presentan las lisas del género Mugil,
también se han abordado los aspectos antes mencionados en otro lugares como
Egipto (Moravec y Libosvarsky, 1975), Portugal (Varela et al., 1981), Francia
(Hubert, 1989), Italia (De Dominis, 1993), China (Lu Junyi et al., 2001) y Brasil
(Ranzani-Paiva y Tavares-Díaz, 2002).
JUSTIFICACIÓN
Aun cuando los registros de Anisakidosis causada por Contracaecum son
escasos, surge el interés por saber si en la población de La Paz, B.C.S. que
consume pescado que ha sido infectado por el nemátodo, se presentan algunos
de los síntomas identificados y relacionados con los de Anisakis, causados en
parte por una reacción alérgica hacia los antígenos del nemátodo. El presente
trabajo presenta información acerca de la composición antigénica de la larva L3
de Contracaecum multipapillatum sensu lato y evalúa su capacidad de inducir
una respuesta inmune específica (inmunogenicidad), información relevante por 1)
los daños producidos por las especies que conforman a la familia Anisakidae en
el hombre (Anisakidosis); 2) la incidencia de Contracaecum en las lisas, siendo
Page 21
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
13
un pez de consumo humano en la localidad; y 3) la posible presencia de
anticuerpos contra un variedad de antígenos que pueden ser identificados por
métodos inmunoquímicos con un extracto somatico total, obtenido de las larvas
L3 de esta especie. Cabe mencionar que éste es el primero de varios estudios
sobre la estructura e inmunogenicidad de los antígenos de la larva L3 de
Contracaecum sp. que ayudarán a establecer el nivel de riesgo a la salud pública
que representa este parásito en productos de consumo humano en la comunidad
de La Paz, B.C.S.
HIPÓTESIS
La larva L3 de Contracaecum sp. en M. cephalus, presenta antígenos que
pueden ser identificables por técnicas inmunoquímicas.
OBJETIVO GENERAL
Estudiar la estructura antigénica de la larva (L3) de Contracacecum sp.
presente en las lisas Mugil cephalus de la Bahía de La Paz.
Objetivos particulares
a) Identificar a nivel especie las larvas (L3) de Contracacecum sp.
b) Obtener el perfil antigénico de la larva (L3) de Contracacecum sp. (por
electroforesis en geles de poliacrilamida)
c) Determinar la inmunogenicidad de los diferentes antígenos de la larva
(L3) de Contracaecum sp. (mediante la inmunización de conejos Nueva Zelandia
bajo condiciones controladas).
Page 22
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
14
d) Estandarización de la técnica de ELISA para la cuantificación de
anticuerpos contra L3 en conejos inmunizados.
e) Identificar los anticuerpos que reconocen antígenos de Contracacecum
sp mediante electroforesis en geles de poliacrilamida e inmunoblotting.
ÁREA DE ESTUDIO
La Bahía de La Paz se localiza en la costa occidental del Golfo de
California entre los 24° 06´y 24° 47´ latitud norte y 110° 18´ y 110° 74´ longitud
oeste (Fig. 4). Tiene una extensión de 2,635 km2, una profundidad máxima de
400m y presenta fondos de arena y fragmentos de carbonato de calcio
(Contreras, 1985; Rueda, 1995; Cruz-Orozco et al., 1996).
La bahía tiene una temperatura mínima superficial de 20° y una máxima de
31°C (Espinoza y Rodríguez, 1987). Cuenta con una precipitación media anual
menor a 200mm, siendo septiembre el mes más lluvioso (>100mm) (Robles Gil-
Mestre, 1998). Está separada del Golfo de California por una península estrecha
y el complejo insular Espíritu Santo-La Partida y presenta una ictiofauna diversa
debido a su gran extensión, profundidad y heterogeneidad ambiental; con un
número total de 522 especies nominales conocidas, siendo el Guachinango
(Lutjanus peru) es la especie más importante en cuanto a captura total, con un
promedio de 760.84 toneladas anuales (Balart et al., 1995; Obeso-Nieblas et al.,
2008; Plomozo-Lugo, 2010).
Page 23
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
15
Figura 4. Área de Estudio. Bahía de La Paz, B. C. S., México.
Page 24
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
16
MATERIAL Y MÉTODOS
Recolecta de material biológico
Hospederos: Se compraron 40 lisas Mugil cephalus de la Bahía de La Paz,
directamente de los pescadores de la marina de la ciudad de La Paz, el mismo
día de su captura y sin eviscerar. Su identificación taxonómica se realizó
mediante el uso de las claves en la Guía FAO para la Identificación de Especies
para los Fines de la Pesca / Pacífico centro-oriental (Fischer et al., 1995).
Nemátodos: Los peces se disecaron en el Laboratorio de Parasitología de
la Universidad Autónoma de Baja California Sur (UABCS) y se realizó una
búsqueda de parásitos por órgano, hasta recolectar 110 larvas de nemátodos de
Contracacecum sp.
Los parámetros ecológicos de prevalencia e intensidad media de infección
por hospedero y por órgano, se obtuvieron de acuerdo al criterio establecido por
Bush et al. (1997).
La prevalencia parasitaria se define como el porcentaje de la población de
hospederos parasitados, respecto a la población de hospederos examinados y se
calcula con la formula:
No. de Hospederos infectados Prevalencia = x 100
No. de Hospederos examinados
La intensidad parasitaria es la cantidad de parásitos encontrados en cada
hospedero revisado y se calcula con la fórmula:
No. de larvas
Intensidad = . No. de Hospederos infectados
Page 25
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
17
Animales de laboratorio: Se adquirieron seis conejos de Nueva Zelandia de
3Kg de peso y se les administró un tratamiento para desparasitarlos,
posteriormente se sometieron a un periodo de cuarentena y aclimatación en el
bioterio del Centro de Investigaciones Biológicas del Noroeste (CIBNOR).
Identificación de las larvas
Se preservaron 10 de las larvas L3 en etanol 95% y se enviaron al Dr.
Robin M. Overstreet de The University of Southern Mississippi - Gulf Coast
Research Laboratory, para realizar la secuenciación de ADN e identificación
taxonómica de las larvas, comparando las secuencias obtenidas con secuencias
conocidas de varias especies del género Contracaecum.
Obtención del extracto crudo de larvas
Los parásitos se lavaron en solución amortiguadora salina de fosfatos
(PBS) para eliminar los restos de tejido de los hospederos, se homogeneizaron
en un homogeneizador de tejidos y se procedió a centrifugar a 6000rpm durante
30 minutos, a una temperatura de 4°C. Se determinó la concentración proteica
del extracto crudo por el método de Bradford (1976) y se almacenó en alícuotas
de 200μl que se conservaron en congelación a -20° C hasta su uso.
Obtención de suero hiperinmune
Se utilizaron cuatro conejos, uno como control y tres a los que se inmunizó
con el extracto crudo obtenido, en un volumen de 2 ml de antígeno en adyuvante
completo de Freund con una concentración final de 100 μg de antígeno por
Page 26
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
18
mililitro, por vía intramuscular cada 14 días hasta cumplir un mes. En el día 16 y
32 se sangraron a los conejos para obtener el suero hiperinmune y el suero
control. A los 37 días los conejos se sacrificaron y se obtuvieron los sueros que
se congelaron hasta su uso en la prueba de inmunoreconocimiento.
Se determinó la concentración de anticuerpos inducidos en conejos, por la
inmunización con los antígenos de Contracaecum sp. por la técnica de ELISA
(Enzyme Linked Immuno-Sorbent Assay) según Yang et al. (1991). En una
microplaca de plástico fueron inmovilizados 10μg (por pozo) de proteínas
somáticas de las larvas, incubándolas toda la noche a 4ºC; después de varios
lavados con PBS-Tween 0.05%, se bloqueó con Albúmina Sérica Bovina al 3%
en PBS. Posteriormente se incubaron con los sueros de los conejos inmunizados
y el conejo control durante dos horas a 37ºC, en dilución seriada y se lavaron
nuevamente con PBS-Tween. Se incubaron 90 minutos a 37ºC con IgG
enlazadas a peroxidasa anti-IgG de conejo, desarrolladas en cabra (Sigma
Inmuno Chemicals / Anti rabbit IgG -whole molecule- peroxidase conjugate).
Después de varios lavados con PBS-Tween, se reveló añadiendo el sustrato y
cromógeno (H2O2, orto-fenil-diamina en solución amortiguadora de citratos pH
5.4) y se incubó en oscuridad a 22ºC durante 30 minutos. Por último, se detuvo la
reacción con H2SO4 1M y se leyó la placa en el cromatógrafo de placas a 492nm.
Análisis electroforético de los antígenos de larvas de Contracaecum sp.
Previo a la realización del análisis, se llevó a cabo el aprendizaje y la
práctica de la técnica en el Centro de Investigación Biomédica de Oriente
(CIBIOR) del Instituto Mexicano del Seguro Social, en Metepec, Puebla.
Page 27
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
19
En el laboratorio de Patología Microbiana del CIBNOR, en La Paz, B.C.S.,
se analizó el extracto crudo de las larvas de Contracaecum sp. por medio de
electroforesis en geles de Poliacrilamida - Dodecil sulfato de sodio (PAGE - SDS)
como describe Laemmli (1970), para ello se usó una cámara de electroforesis
Protean II (gel grande) y una Mini Protean II (gel chico).
Los geles consistieron en gel concentrador al 4% y gel separador al 20%.
Se utilizó solución amortiguadora para electrodos (Tris 25mM, glicina 192mM,
SDS 1%, pH 8.3) y solución amortiguadora para muestras (Tris-HCl 50mM pH 8.6
conteniendo SDS 2% glicerol 20%, 2-mercaptoetanol 2% y azul de bromofenol
0.02%) para disolver las muestras 1:1 (18μl-muestra / 18μl-solución
amortiguadora para muestra por pozo en geles chicos y 40μl-muestra / 40μl -
solución amortiguadora para muestra por pozo en geles grandes). Las
electroforesis se realizaron a 100 V durante 2 horas. Se corrieron marcadores de
pesos moleculares “BIO-RAD SDS-PAGE Molecular weight standards” de rango
amplio (200,000 - 6,500 kDa) y rango bajo (97,400 - 14,400 kDa) (Tabla 1) junto
con las muestras.
Tabla 1. Proteínas y sus respectivos pesos moleculares del estándar “BIO-RAD SDS-PAGE Molecular weight standards” (Cat-Num 161-0304)
Proteína Peso molecular (kDa)
Fosforilasa b 97.4
Albúmina Sérica Bovina 66.2
Ovoalbúmina 45
Anhidrasa Carbónica 31
Inhibidor de Tripsina de soya 21.5
Lisozima 14.4
Page 28
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
20
Inmunoblotting de los antígenos de larvas de Contracaecum sp.
Previo a la realización del análisis, se llevó a cabo el aprendizaje y la
práctica de la técnica en el CIBIOR del Instituto Mexicano del Seguro Social.
Se realizó un análisis de inmunoreconocimiento mediante una electro-
transferencia de las proteínas del extracto crudo previamente separado mediante
PAGE-SDS, a membranas de polifloruro de vinilideno (Immobilon - P Millipore)
con poros de 0.4 µm utilizando para ello una cámara de transferencia semi-seca
(Bio-Rad Trans-Blot SD Semi-Dry Transfer Cell) durante 30 minutos a 20V,
utilizando una solución amortiguadora de transferencia (Tris 25mM, glicina
192mM, metanol 20%, pH 8.3). Las membranas se bloquearon con leche
descremada al 5% a 4°C durante toda la noche; posteriormente se lavaron tres
veces con solución de lavado (PBS-BSA 5% Tween20 2%).
Se separó una muestra de cada membrana para ser incubada con suero
de conejo no inmunizado (control), diluido en PBS 1:60 durante 2 horas con
agitación; el resto de las muestras se incubaron con suero de conejo anti-
Contracaecum diluido en PBS al 1:60 por 2 horas con agitación a temperatura
ambiente. Todas las muestras se lavaron tres veces más, con solución de lavado
y se incubaron por 1 hora con un anticuerpo anti-IgG de conejo acoplado a
peroxidasa desarrollado en cabra, diluido 1:1000 en PBS-Tween20 conteniendo
BSA al 1%, a temperatura ambiente. Para visualizar las bandas, se lavaron las
membranas con PBS-Tween 20 y se agregó el sustrato (PBS, H2O2 al 0.006% en
metanol conteniendo 0.03% de 4-cloro-1-naftol); finalmente se detuvo la reacción
con varios lavados de agua destilada (Fig. 5).
Page 29
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
21
Figura 5. Diagrama de flujo de los principales pasos de la metodología realizada.
Compra
Compra
Gulf Coast Research Laboratory
Cuarentena y aclimatación
Preparación de
Análisis por
Sangrado Transferencia
a membrana
Inoculación
Inyección intramuscular
Obtención
Cálculo de Pesos moleculares
ELISA
Recolecta de Material Biológico Mugil cephalus Nemátodos
Conejos
Electroforesis en geles de poliacrilamida
SDS-PAGE Inmunización
Inmunoblotting
Suero hiperinmune
Perfil Proteico (Antígenos)
Antígenos reconocidos por IgG’s de conejo
Identificación taxonómica molecular
Extracto crudo
METODOLOGÍA
Page 30
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
22
RESULTADOS
Se revisaron 40 lisas presentando un 100% de prevalencia parasitaria y se
obtuvieron un total de 521 nemátodos en su tercer estadio larvario (Tabla 2).
Los órganos en los que se encontraron los parásitos fueron, en orden
descendente de abundancia: el hígado, serosa del estómago-intestino, riñón,
mesenterio de la cavidad celómica (M.C.C) y la musculatura parietal (filete)
(Tabla 2).
Tabla 2. Prevalencia e intensidad parasitaria de C. multipapillatum s.l. (L3) por hospedero y en los diferentes órganos de Mugil cephalus.
No. de Larvas Prevalencia Intensidad
Hospederos 521 100% 13
Hígado 232 100% 5.8
Serosa Estómago-Intestino 171 95% 4.5
Riñón 103 80% 3.2
M.C.C. 12 17.5% 1.7
Músculo 3 5% 1.5
Identificación taxonómica de hospederos y nemátodos
Los peces recolectados fueron identificados como Mugil cephalus
(Linnaeus 1758), basándose en su coloración característica y a la presencia de:
un recubrimiento adiposo extendido sobre el iris; el número de escamas
longitudinales (38 a 42); la segunda aleta dorsal con 9 radios y la aleta anal con 3
espinas y 8 radios.
Page 31
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
23
Las larvas de nemátodos extraídas de M. cephalus corresponden a la fase
3 (L3) del género Contracaecum. Dentro de sus características morfológicas se
observan: tres labios y un diente cuticular; un ventrículo pequeño y esférico;
apéndice ventricular dirigido hacia el extremo posterior y un ciego intestinal, más
grande que el apéndice ventricular. No fue posible identificar a nivel especifico los
especímenes obtenidos, pero el análisis molecular indica que pertenecen al
complejo multiespecífico de Contracaecum multipapillatum, taxón dentro del que
se han descrito, hasta el momento, cuatro especies. Los especímenes de este
estudio deben ser referidos como Contracaecum multipapillatum sensu lato.
Análisis electroforético de los antígenos de larvas de C. multipapillatum s. l.
El extracto crudo somático de las larvas L3 de C. multipapillatum s. l.
presentó una concentración de 80 μg/μl·prot. En el extracto crudo se identificaron
54 proteínas por el método de electroforesis SDS-PAGE (Fig. 6). Las proteínas
en el extracto se encuentran en un intervalo de pesos moleculares de 1.2417a
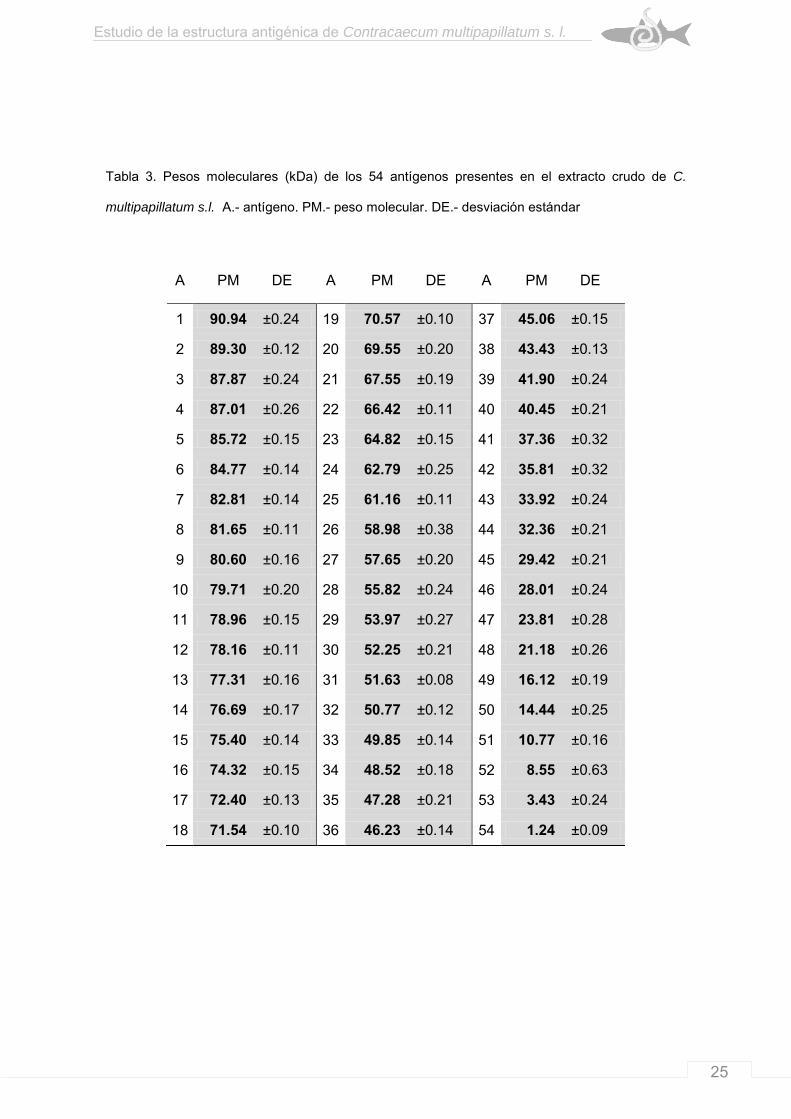
90.9431 kDa (Tabla 3).
Page 32
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
24
Figura 6. Perfiles proteicos del extracto crudo de C. multipapillatum s.l. en geles de electroforesis
SDS-PAGE. A1 Marcadores de peso molecular en gel chico. A2 Antígenos de C. multipapillatum
s.l. en gel chico. B1 Marcadores de peso molecular en gel grande. B2 Antígenos de C.
multipapillatum s.l. en gel grande.
Page 33
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
25
Tabla 3. Pesos moleculares (kDa) de los 54 antígenos presentes en el extracto crudo de C.
multipapillatum s.l. A.- antígeno. PM.- peso molecular. DE.- desviación estándar
A PM DE A PM DE A PM DE
1 90.94 ±0.24 19 70.57 ±0.10 37 45.06 ±0.15
2 89.30 ±0.12 20 69.55 ±0.20 38 43.43 ±0.13
3 87.87 ±0.24 21 67.55 ±0.19 39 41.90 ±0.24
4 87.01 ±0.26 22 66.42 ±0.11 40 40.45 ±0.21
5 85.72 ±0.15 23 64.82 ±0.15 41 37.36 ±0.32
6 84.77 ±0.14 24 62.79 ±0.25 42 35.81 ±0.32
7 82.81 ±0.14 25 61.16 ±0.11 43 33.92 ±0.24
8 81.65 ±0.11 26 58.98 ±0.38 44 32.36 ±0.21
9 80.60 ±0.16 27 57.65 ±0.20 45 29.42 ±0.21
10 79.71 ±0.20 28 55.82 ±0.24 46 28.01 ±0.24
11 78.96 ±0.15 29 53.97 ±0.27 47 23.81 ±0.28
12 78.16 ±0.11 30 52.25 ±0.21 48 21.18 ±0.26
13 77.31 ±0.16 31 51.63 ±0.08 49 16.12 ±0.19
14 76.69 ±0.17 32 50.77 ±0.12 50 14.44 ±0.25
15 75.40 ±0.14 33 49.85 ±0.14 51 10.77 ±0.16
16 74.32 ±0.15 34 48.52 ±0.18 52 8.55 ±0.63
17 72.40 ±0.13 35 47.28 ±0.21 53 3.43 ±0.24
18 71.54 ±0.10 36 46.23 ±0.14 54 1.24 ±0.09
Page 34
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
26
Obtención de suero hiperinmune e inmunoreconocimiento de los antígenos
El suero analizado se obtuvo de tres conejos inmunizados con extracto
crudo de C. multipapillatum s.l., ya que uno de ellos falleció antes del primer
sangrado. La prueba de ELISA practicada, mostró que el suero extraído del
sangrado del día 16 después de la inmunización presentó niveles bajos (no
cuantificables) de inmunoglobulinas G (IgG) anti - Contracaecum, mientras que
los sueros obtenidos de los sangrados de los días 32 y 37 presentaron 0.6μg/μl y
0.57μg/μl respectivamente.
En el suero del conejo control existen anticuerpos tipo IgG que reconocen
14 de los 54 antígenos presentes en el extracto crudo de C. multipapillatum s.l.
(Tabla 3, columna A). El suero de los conejos inmunizados presentó anticuerpos
capaces de reconocer 33 de esos 54 antígenos (Fig. 7) por lo que estos
resultados demuestran que el sistema inmune de los conejos utilizados en este
estudio puede producir IgG’s capaces de reconocer 19 de los antígenos con
aparente especificidad hacia C. multipapillatum s.l. (pesos moleculares entre
90.94 y 8.55 kDa) (Tabla 4, columna B*)
Page 35
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
27
Figura 7. Antígenos de C. multipapillatum s.l. reconocidos por IgG’s presentes en suero de conejo.
A. Antígenos reconocidos por suero de conejo control (14). B. Antígenos reconocidos por suero
de conejo inmunizado (33)
Page 36
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
28
Tabla 4. Antígenos de C. multipapillatum s.l. reconocidos por suero de conejo. A. Pesos
moleculares (kDa) de los antígenos reconocidos por suero de conejo control (14). B. Pesos
moleculares (kDa) de los antígenos reconocidos por suero de conejos inmunizados (33).
A B - 90.94* - 87.87* - 87.01* - 82.81* - 81.65* - 80.60* - 79.71*
78.96 78.96 78.16 78.16
- 77.31* - 75.40* - 72.40*
71.54 71.54 - 69.55*
67.55 67.55 66.42 69.55
- 69.55* 62.79 62.79 58.98 58.98 53.97 53.97
- 52.25* - 50.77* - 49.85*
46.23 46.23 43.43 43.43 40.45 40.45 35.81 35.81
- 33.92* - 32.36*
23.81 23.81 16.12 16.12
- 10.77* - 8.55*
*Pesos moleculares de los antígenos con aparente especificidad, reconocidos solo por el suero
de los conejos inmunizados (19).
Page 37
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
29
DISCUSIÓN
Identificación taxonómica.
En el área de la epidemiología es importante conocer con claridad qué
especies son las causantes de afecciones en el humano. Debido a que algunas
de las características diagnósticas utilizadas en la identificación morfológica de
las especies carecen de valor interespecífico, el uso de herramientas moleculares
para una correcta identificación de los parásitos ha cobrado gran importancia. Se
han descrito numerosas especies nuevas de anisákidos en las últimas dos
décadas, debido a la hallazgo de varias especies hermanas, con pozas genéticas
aisladas reproductivamente pero con morfología muy similar. La falta de
diferenciación puede deberse a que estos parásitos se ven afectados por factores
de selección muy similares, causando la conservación de las características
morfológicas, además de que muchas de estas características parecen ser
altamente homoplásicas o representan estados morfológicos ancestrales
(Mattiucci y Nascetti, 2008).
Los análisis realizados en The University of Southern Mississippi - Gulf
Coast Research Laboratory por el Dr. Robin Overstreet a las larvas de nematodo
obtenidas de Mugil cephalus, indican que éstas pertenecen a una especie
genéticamente muy cercana a las especies del complejo de Contracaecum
multipapillatum, pero no fue posible identificar el nivel específico al que
pertenecen. Es necesario llevar a cabo nuevos análisis que confirmen si estas
larvas pertenecen a una de las 4 especies que están actualmente dentro del
complejo o si es una especie nueva. Por este motivo la manera indicada para
referirse a los especímenes encontrados en las lisas de la Bahía de La Paz es
Page 38
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
30
Contracaecum multipapillatum sensu lato, que indica que se trata de una especie
dentro del taxón Contracaecum multipapillatum, cuyo volumen es más amplio que
el descrito en este trabajo en particular.
Parasitismo por Contracaecum
Ecológicamente los parásitos son una parte esencial de la comunidad
acuática y la mayoría de los individuos de las poblaciones de peces, silvestres o
en cultivo, están infectados con uno o más parásitos (Al-Zubaidy, 2009).
Al parecer, los parásitos anisákidos se acumulan durante el tiempo de vida
de los peces; la prevalencia e intensidad de infección por nemátodos aumenta a
lo largo de la ontogenia, conforme crece el pez. De acuerdo con Moravec et al.
(1995) para que un parásito se albergue en un hospedero, éste necesita ser
abundante en el medio. Las posibilidades de infección aumentan debido al
periodo de exposición a las fases infectivas del nemátodo; al aumento en la
cantidad de comida que ingiere; a la acumulación de parásitos a lo largo de la
cadena alimenticia, en particular en el caso de endoparásitos y probablemente
también al cambio de dieta durante su vida (Szalai y Dick, 1990; Aho y Bush,
1993; Bergmann y Motta, 2004; Al-Zubaidy, 2009). En el caso de M. cephalus
Valles-Ríos et al. (2000) han reconocido una correlación positiva entre el tamaño
del pez y la intensidad de infección de Contracaecum spp. y otros parásitos.
Una de las causas de las infecciones en peces son los cambios de
factores bióticos o abióticos en el ambiente. Varias condiciones patológicas como
las alergias y reacciones anafilácticas ocurren en personas que se alimentan de
peces que habitan en cuerpos de agua contaminados, en los que aumenta la
Page 39
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
31
concentración de plancton cuyos organismos pueden actuar como hospederos
intermediarios o paraténicos de helmintos, que a su vez, podría incrementar el
número de infecciones en el siguiente nivel de la cadena trófica (Mattiucci y
Nascetti, 2008; Al-Zubaidy, 2009).
En el presente estudio se obtuvo una prevalencia parasitaria por
Contracaecum multipapillatum (sensu lato) de 100% en M. cephalus obtenidas
en los meses de octubre y noviembre, similar a lo encontrado por Gómez del
Prado-Rosas et al. (2008) en M. cephalus parasitadas por Contracaecum sp. en
tres localidades dentro de la Bahía de La Paz (San Juan de la Costa, El Comitán
y Punta Prieta) durante el verano de 1995 y Valles Vega et al. (2010) en M.
curema de la Bahía de La Paz, mientras que la prevalencia de larvas de C.
multipapillatum en M. cephalus de Baja California obtenidas por Valles-Ríos et al.
(2000) fue de 30% y en otros estudios como el de Iannacone y Alvariño (2009) en
Perú, llegan a ser tan bajas como 2.7 %. Esto sugiere que algún factor,
probablemente la eutrofización, en la Bahía de la Paz, conduce a la infección de
un alto porcentaje de las lisas en el área.
Por muchos años la Ensenada de La Paz recibió la descarga directa de
aguas residuales de la ciudad y varios autores registraron altos niveles de
contaminación en las décadas de 1970 y 1980 (Martínez Díaz y Hernández
Rivas, 2010). En la actualidad, del total de aguas residuales tratadas en lagunas
de estabilización, aproximadamente un 35% se utiliza en riego agrícola, el resto
se vierte en la Ensenada de La Paz, por lo que grandes cantidades de materia
orgánica y bacterias son descargadas en la ensenada (Camacho, 1996; Martínez
Díaz y Hernández Rivas, 2010).
Page 40
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
32
Olivero et al. (2006) encontraron diferencias dramáticas en las parasitosis
por nemátodos en el pez de agua dulce Halopias malabaricus de Colombia,
registrando mayores prevalencia e intensidad de parasitismo en los peces que
habitan algunos cuerpos de agua que mostraron más contaminación. Por otro
lado, Al Zubaidy (2009) menciona que, como la diversidad de peces disminuye en
ambientes acuáticos contaminados, el problema de las infecciones por helmintos
en humanos, adquiridas por el consumo de pescado, se acrecienta, ya que
solamente pocas especies están disponibles (Sakanari y McKerrow, 1989).
A pesar de que la mayor parte del año, la Ensenada de La Paz posee las
características adecuadas para uso recreativo y pesca, es importante considerar
los efectos naturales como el aumento en la precipitación pluvial y la insuficiencia
de los sistemas de recolección de agua del alcantarillado en el Municipio de La
Paz, que provocan un aumento en la contaminación en algunas temporadas del
año como en la época de huracanes (Martínez Díaz y Hernández Rivas, 2010).
Por el contrario, en la Bahía de La Paz no se tienen registros de
contaminación por aguas residuales, sin embargo, debido al sistema de
circulación de corrientes de la Ensenada de La Paz, las aguas residuales que se
llegan a verter en ésta, terminan finalmente en la bahía. Además de la actual
tendencia al aumento en la infraestructura turística proyectada en todo el margen
de la ensenada y la parte sur de la Bahía de La Paz, que incluye la instalación de
zonas habitacionales, turísticas y recreativas en áreas naturales (Martínez Díaz y
Hernández Rivas, 2010).
Por lo tanto, serían de utilidad estudios que esclarezcan si existe una
relación entre los niveles de contaminación biológica en la Ensenada y la Bahía
Page 41
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
33
de La Paz y la presencia de C. multipapillatum s. l. y otros helmintos parásitos en
el ecosistema. Es posible que el ciclo de vida de los nemátodos anisákidos esté
relacionado con la demografía de sus hospederos en un área particular, debido a
que la culminación de sus ciclos de vida tan complejos requiere estabilidad en la
red trófica, ya que interviene en distintos niveles de ésta. Varios estudios indican
que existen efectos e interacciones significativas entre los niveles de parasitismo y
la presencia y concentracion de contaminantes y otros factores de estres en el
en el ambiente que pueden causar la disminución de las poblaciones hospederos
por perturbaciones en el ambiente como la sobrepesca, captura incidental, etc.,
que se manifiestan también en la disminución de las poblaciones de anisákidos
endoparásitos (Vidal - Martínez et al., 1994 y 2009; Coscia y Oreste, 1998;
Mattiucci et al., 2008; Mattiucci y Nascetti, 2008; Al-Zubaidy, 2009; Martínez Díaz
y Hernández Rivas, 2010).
Al peligro potencial que representa la alta prevalencia de Contracaecum
multipapillatum s.l. y otras posibles especies del género en distintos peces de la
Bahía de La Paz, se suma el hecho de que la temperatura del agua en que se
desarrollan los parásitos puede influir en su capacidad de infectar mamíferos y
por lo tanto al humano (Vidal-Martínez et al., 1994).
Ko, (1976, 1977) demostró que una mayor cantidad de infecciones de
distintos mamíferos con el nemátodo Echinocephalus sinensis obtenido de ostión,
se presentaron con especímenes recolectados en ciertos meses con mayores
temperaturas y con especímenes aclimatados a 28 y 33 ºC, temperaturas
Page 42
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
34
comunes en las localidades donde Contracaecum multipapillatum es abundante,
mientras que pocas o ninguna infección se presentaron con los nemátodos
aclimatados a 5, 15 y 20 - 24 ºC. Por este motivo también resultan de interés
estudios que exploren las variaciones en la capacidad infectiva de los parásitos
que pueden amenazar la salud pública en la localidad en distintas temporadas del
año, ya que en la Bahía de la Paz existen variaciones significativas en las
temperaturas superficiales del agua, las más altas se registran de julio a octubre
(30 - 32 ºC) y las más bajas de enero a marzo (20 - 22 ºC) (Vidal-Martínez et al.,
1994; López et al., 2001).
Muchos parásitos pueden alterar el funcionamiento de los órganos vitales
de los peces de diversas maneras: pueden afectar la actividad del corazón, en el
intestino pueden inhibir la actividad digestiva y el metabolismo de vitaminas y
azúcar, por lo tanto, de manera indirecta afectan el crecimiento; en el hígado
afectan el metabolismo del glucógeno, en gónadas y cavidad celómica provocan
la reducción del número de huevos o la castración (Rhode, 1993).
En el caso de las lisas capturadas en la Bahía de La Paz, su aspecto
externo no muestra signos de enfermedad, a pesar de la alta intensidad de la
parasitosis en órganos viscerales por larvas del tercer estadio de C.
multipapillatum s. l. El único órgano que, en ocasiones, presenta lesiones
evidentes a simple vista, al abrir la cavidad visceral de los peces, es el hígado,
por lo que su valor y comercialización no disminuye. Por esta razón, aumentan
los riesgos de infección con este parásito o de presentar una reacción alérgica
hacia los antígenos del gusano o los presentes en la carne del pescado que
estuvo en contacto con el parásito (Al-Zubaidy, 2009).
Page 43
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
35
Narciso et al. (2010) encontraron en M. cephalus a la venta dentro del área
metropolitana de Monterrey, Nuevo León, México, que los sitios con mayor
número de larvas de Contracaecum sp. son el hígado, seguido por el músculo
(filete), pero no encontró ningún nemátodo en el intestino, lo que hace suponer
que al ser más largo el periodo que pasa el pescado sin ser eviscerado al
transportarse a zonas del país lejanas a la costa, se promueve la migración de
las larvas alojadas en intestino y otros órganos hacia la musculatura, como
sucede con varios anisákidos.
De las 40 lisas recolectadas en el presente trabajo, solamente en dos se
encontraron larvas parasitando el músculo, por ello, se recomienda eviscerar el
pescado inmediatamente después de su captura, con el fin de reducir la infección
en músculo y no consumir pescado que lleve más de un día sin haber sido
eviscerado.
Aun así, probablemente no existe una preparación al momento de cocinar
que elimine por completo el riesgo a la salud humana cuando se consume
pescado que fue parasitado, en parte, porque ningún método garantiza al
consumidor, protección contra una posible reacción alérgica causada por la
ingestión de antígenos del parásito; en especial cuando algunos alérgenos de
larvas en tercer estadio (Anisakis simplex) han mostrado resistencia a
temperaturas elevadas en pruebas in vitro (Buendía E., 2000; Skirnisson, 2006).
Page 44
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
36
Estructura antigénica e inmunogenicidad de
Contracaecum multipapillatum s. l.
El reconocimiento de antígenos de nemátodos aumenta nuestra capacidad
de entendimiento de los mecanismos de defensa desencadenados por el
hospedero durante la infección, ya que la respuesta inmune es la responsable de
muchos de los cambios patológicos que se presentan. Esto ha dado lugar al
desarrollo de métodos inmunodiagnósticos específicos, de medicamentos y
vacunas (Perec y Okulewicz, 2006; MacDonald et al. 2002).
En la literatura se menciona que la primera respuesta del sistema inmune
hacia antígenos, “respuesta primaria”, ocurre después de una primera inoculación
con presencia de inmunoglobulinas IgM principalmente y éstas deben alcanzar su
concentración óptima entre los días 10 y 15 después de la inoculación. Después
de este periodo, una segunda inoculación genera la “respuesta secundaria” en la
que, pocos días después, los linfocitos B producen las inmunoglobulinas IgG.
En el presente trabajo, el suero de los conejos inmunizados con proteínas
somáticas de las larvas L3 obtenido del sangrado del día 32, es decir, cuatro días
después de la segunda inmunización, fue el que presentó una mayor
concentración de IgG Anti - C. multipapillatum s. l., sin que existieran diferencias
en cuanto a la cantidad y peso molecular de los antígenos que son capaces de
reconocer, respecto a los sueros obtenidos el día 37. La metodología empleada
es similar a la registrada por Escalante et al. (2005) y López-Arellano et al.
(2005), que estudiaron la inmunogenicidad de los antígenos de nemátodos
Ascaris suum (parásito de roedores y humanos) y Haemonchus contortus
(parásito de rumiantes), utilizando conejos con un protocolo de inmunización con
Page 45
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
37
intervalos de 7 o 15 días, inoculando proteínas somáticas junto con adyuvante
completo de Freund, los sueros hiperinmunes que utilizaron, se obtuvieron a
partir de la “respuesta secundaria” del sistema inmune de los conejos. Después
de la reinmunización, la “respuesta secundaria” alcanza niveles mayores de
anticuerpos IgG, que descienden lentamente en los siguientes dos o tres meses
(Calderón-Tinoco, 1984; Tizard, 2002;).
Sin embargo, la sensibilidad y exactitud de las pruebas inmunológicas en
modelos animales se ven afectadas cuando existen factores que modifiquen la
respuesta inmune del sujeto (Perec y Okulewicz, 2006). De los seis conejos
adquiridos que se sometieron al tratamiento de desparasitación y los periodos de
cuarentena y aclimatación en el bioterio del CIBNOR, sobrevivieron cinco, de los
cuales uno sufrió una fractura expuesta en el quinto día después de la primera
inoculación y falleció en el séptimo día sin que se determinara la causa especifica
de la muerte. Se realizó una extracción de sangre de este conejo para obtener
suero, con el que se realizó la prueba de ELISA para conocer la concentración de
IgG’s Antí - C. multipapillatum s.l. tras los siete días de inoculación. Debido a que,
probablemente, en este conejo se produjo un efecto inmunosupresor tras la
fractura y pérdida de sangre y falleció solo siete días después de la primera
inoculación, las concentraciones de IgG’s Antí - C.multipapillatum s. l. presentes
en su suero no fueron identificadas por el método de ELISA.
El reconocimiento de antígenos de helmintos parásitos por parte del
sistema inmune, puede verse afectado como consecuencia a que éstos se han
adaptado a suprimir, contrarrestar o evitar el sistema inmunitario de sus
hospederos. A través de la secreción de hormonas y enzimas pueden evitar la
Page 46
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
38
producción o favorecer la degradación de inmunoglobulinas, además, pueden
evadir la acción proteolítica de las células defensivas con inhibidores de
proteasas o antioxidantes (Tizard, 2002).
Las larvas del nemátodo Toxocara canis, al ser expuestas in vitro a
eosinófilos del hospedero, simplemente se desprenden de la cubierta larvaria
junto con las células adheridas, evitando su ataque. Sin embargo, en la respuesta
inmune de los hospederos naturales de algunos parásitos, se ha observado que a
pesar de que no siempre reconocen los antígenos de las superficies corporales
de los helmintos, si son capaces de reconocer los antígenos que no están
comúnmente en contacto directo con ellos como los productos de secreción o
proteínas del tracto digestivo (Tizard, 2002).
Por ello, es importante conocer el total de los antígenos somáticos de un
parásito, capaces de generar una respuesta inmune en mamíferos, para el
desarrollo de métodos de diagnóstico, fármacos y/o vacunas eficientes.
En el extracto crudo somático total de larvas L3 de C. multipapillatum s. l.
se identificaron 54 proteínas de las cuales 14 fueron reconocidas por las IgG en
el suero del conejo control con pesos moleculares entre los 23.81 y 78.96 kDa,
mientras que el suero de los conejos inmunizados reconoció 33 proteínas. Esto
quiere decir que, los conejos inmunizados produjeron IgG’s por la presencia de
los antígenos C. multipapillatum s. l. para 19 antígenos. Es decir, produjo 19
IgG’s anti - C. multipapillatum s. l. sin que esto quiera decir que son específicas
para los antígenos de esta especie, ya que hace falta realizar pruebas que
descarten posibles reacciones cruzadas con antígenos de otros nemátodos u
otros helmintos.
Page 47
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
39
En contraste, Perec y Okulewicz (2006) obtuvieron 47 proteínas somáticas
del nemátodo Syphacia obvelata, 11 de éstas con pesos moleculares mayores a
las proteínas somáticas de C. multipapilltum s. l. Debido a que uno de los factores
que determina la inmunogenicidad de los antígenos es su complejidad y peso
molecular, es de suponerse que las proteínas somáticas de S. obvelata tienen la
capacidad de estimular en mayor medida la producción de anticuerpos. Del total
de proteínas de S. obvelata 72 % (34) fueron reconocidas por IgG’s de ratones de
laboratorio de la cepa BALB/c, en contraste, las IgG’s de los conejos inmunizados
de este estudio sólo reconocen un 61% (33).
A pesar de que los resultados indican que C. multipapillatum s. l. posee
menos proteínas inmunogénicas que S. obvelata, el hecho de que en el estudio
de Perec y Okulewicz (2006) no presentan una comparación entre la cantidad de
antígenos reconocidos por IgG’s de ratones infectados y los reconocidos por
IgG’s de ratones no infectados, no nos permite hacer una comparación clara de
cuántos de los antígenos de cada nemátodo son reconocidas por anticuerpos
producidos específicamente por la exposición al parásito (o su extracto crudo) y
cuáles están presentes de manera natural, antes de que sea estimulado el
sistema inmune de los sujetos de estudio.
Coscia y Oreste (1998) observaron solamente 14 bandas
correspondientes a las proteínas de Contracaecum osculatum con pesos
moleculares entre los 21 y 200 kDa. En pruebas de inmunoreconocimiento
encontraron que el pez teleósteo Chionodraco hamatus presentó
inmunoglobulinas que reconocen 21% (3) de estas proteínas con pesos
moleculares de 47, 63 y 91 kDa. Como se mencionó anteriormente los
Page 48
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
40
hospederos naturales de algunos helmintos no son capaces de reconocer varios
de sus antígenos, en consecuencia a la adaptación de éstos al medio que
presenta el hospedero y su sistema inmune. Esto podría explicar el bajo
porcentaje de proteínas reconocidas por los peces, hospederos intermediarios
naturales de las larvas de Contracaecum spp. comparado con el porcentaje de
proteínas reconocidas por el sistema inmune de los conejos del presente trabajo,
que no son hospederos naturales de las especies de este género.
Como parte del presente estudio se tenía contemplado la realización de
pruebas de inmunoreconocimiento de los antígenos de C. multipapillatum s. l. en
el suero de sus hospederos intermediarios naturales, M. cephalus, sin embargo,
debido a que el procesamiento de las muestras del suero de los peces no se
realizó de manera adecuada, no fue posible utilizarlas. Estos análisis nos pueden
dar más información y ser un punto de referencia acerca de qué proteínas de las
distintas especies del género Contracaecum presentan mayor inmunogenicidad.
No obstante, para hacer una comparación apropiada con estudios como el
de Coscia y Oreste (1998), es necesaria la homogenización de protocolos que
permitan una mayor eficacia para la obtención de antígenos. A pesar de que
analizaron proteínas de un nemátodo del mismo género al estudiado en el
presente trabajo, el número de proteínas obtenidas en cada uno, es muy distinto.
Los resultados obtenidos a partir de Contracaecum multipapillatum sensu lato son
más similares a los descritos por Perec y Okulewicz (2006) en Syphacia obvelata,
miembro de una familia distinta.
Page 49
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
41
Futuro de la línea de investigación
En próximos trabajos se buscará esclarecer si los nemátodos encontrados
en M. cephalus corresponden a alguna de las especies ya descritas dentro del
complejo de Contracaecum multipapillatum, o si se trata de una nueva especie,
para ello será necesario conocer las características morfológicas de los adultos y
qué aves actúan como hospederos definitivos de esta especie. Esto proveerá un
mejor conocimiento de su ciclo de vida, que como ya se mencionó, también
puede ayudar a entender los niveles de infección de la especie en el área.
Además, en los estudios que deriven de los resultados presentados en el
presente trabajo será necesario realizar experimentos en búsqueda de
anticuerpos con mayor afinidad e inmunogenicidad, en modelos animales
mamíferos y sus hospederos peces y aves. Será necesario llevar a cabo un
estudio prospectivo para determinar si existen y con qué frecuencia se presentan
las infecciones o reacciones alérgicas provocadas por las larvas de
C. multipapillatum s. l., en pacientes con alguno o varios de los síntomas
relacionados con la anisakiasis después de haber consumido pescado de la
Bahía de La Paz. Para ello también es recomendable analizar las posibles
reacciones cruzadas de los antígenos aquí mostrados, con antígenos de otros
parásitos presentes en los peces de consumo en la localidad, para determinar
cuáles pueden ser utilizados en pruebas diagnósticas más exactas.
Page 50
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
42
CONCLUSIONES
• Las larvas L3 encontradas en Mugil cephalus de la Bahía de La Paz se
ubican taxonómicamente dentro del complejo de Contracaecum multipapillatum,
que incluye a C. gibsoni, C. overstreeti, C. multipapillatum sp. “C” y C.
multipapillatum sp. “D”. Pero es necesario determinar si corresponden a alguna
de estas cuatro especies o se trata de una especie no descrita aún.
• Las lisas M. cephalus de la Bahía de La Paz, parasitadas por larvas de C.
multipapillatum s. l. presentan prevalencias e intensidades inusualmente altas,
siendo mayor la infestación de hígado e intestino y menos común en la
musculatura parietal de peces recién capturados.
• La presencia de larvas de C. multipapillatum s. l. con altas prevalencia e
intensidad en M. cephalus no afecta la comercialización de este pez, por lo tanto,
el peligro potencial de infección o de presentar una reacción alérgica, está
presente en las personas que lo consumen.
• En las larvas L3 de C. multipapillatum s. l. es posible identificar 54
proteínas con pesos moleculares entre 1.24 y 90.94 kDa por medio de
electroforesis SDS en gel de poliacrilamida.
• De las proteínas de C. multipapillatum s. l., 61% (33) son capaces de
inducir la producción de IgG’s en conejos. De estos 33 antígenos, 19 generan la
producción de IgG’s anti - C. multipapillatum s. l.
Page 51
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
43
RECOMENDACIONES
Al estudiar los efectos de una determinada especie parásita en el
ecosistema o en la salud pública es de gran utilidad acceder al tema de manera
interdisciplinaria. En primera instancia se debe identificar taxonómicamente de
manera concluyente a la o las especies que son el objeto de estudio, para ello, el
uso de herramientas de biología molecular han probado ser de gran utilidad al
estudiar helmintos.
A su vez, para entender la dinámica de las poblaciones de parásitos dentro
de sus hospederos y en el ecosistema, el estudio de las estrategias que
desarrollan ambos organismos para coexistir, incluyendo la respuesta inmune del
hospedero, podrían ser de gran ayuda para explicar el porqué de temas como: la
capacidad de los helmintos de invadir nuevos tejidos o nuevos hospederos; la
habilidad del hospedero para evitar infestaciones masivas al estar expuesto
constantemente a las fases infectivas de determinados helmintos; la proporción
de parásitos adultos y juveniles dentro de un mismo hospedero, así como la
presencia o ausencia de diferentes especies de parásitos y otros aspectos de la
ecología y epidemiología de los helmintos y sus hospederos, incluido el humano.
También se debe analizar la posibilidad de utilizar el estudio de la ecología
y variabilidad genética de las poblaciones de C. multipapillatum s. l. y otros
parásitos en el área, como parámetros del estado de salud del ambiente, debido
a la estrecha relación que existe entre el ciclo de vida de algunos parásitos con
organismos en diversos nichos de un ecosistema. Probablemente Contracaecum
multipapillatum s. l. puede ser utilizado como indicador de la salud de las
poblaciones de peces y dentro de la Bahía de La Paz, ya que existen datos que
Page 52
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
44
indican que la alta prevalencia de infección, puede estar influenciada por la
eutrofización en la Ensenada y Bahía de La Paz, debida a las descargas de
materia orgánica provenientes del drenaje de la ciudad o provocadas por eventos
naturales como huracanes.
Respecto a la amenaza que puede representar C. multipapillatum s. l. a la
salud pública de la región, es recomendable, con el fin de disminuir la
probabilidad de ingerir accidentalmente las larvas o que sus antígenos estén
presentes en la carne del pescado, evitar el consumo de pescado que no ha sido
eviscerado de manera oportuna después de su captura. Pero se deberá exhortar
también a los comerciantes iniciales, los pescadores, y a los intermediarios, a
adoptar medidas de prevención, eviscerando el pescado que comercializan, lo
más pronto que les sea posible.
Por otra parte se debe promover en los grandes establecimientos y
empresas con mayores recursos económicos, la implementación de medidas
como congelar a una temperatura de -20°C durante más de 24 horas todo el
pescado que debe comercializarse crudo, actualmente obligatorio en la
Comunidad Económica Europea.
Se recomienda alertar a las autoridades sanitarias de las circunstancias en
las que se encuentran las lisas de venta en el área, y el posible peligro al que se
exponen sus consumidores, para que tomen medidas precautorias y participen en
las investigaciones que determinen la magnitud del riesgo que representa la
especie objeto de este trabajo y otras especies del mismo género posiblemente
presentes en los organismos de los que se obtienen productos de consumo
humano.
Page 53
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
45
TRABAJOS FUTUROS
Actualmente se trabaja en la secuenciación del ITS-1, ITS-2 (rADN) y el
COx2 (mtADN) de las larvas de Contracaecum provenientes de Mugil cephalus y
Mugil curema encontrándose hasta el momento, que estos nemátodos presentes
en ambas, pertenecen al complejo de Contracaecum multipapillatum. Se busca
determinar si se tratan de una nueva especie dentro de este complejo o si
pertenecen a una de las cuatro especies ya descritas. También está en proceso
el análisis de las mismas secuencias en adultos obtenidos de Pelecannus
occidentalis.
Es posible estudiar si la capacidad infectiva de C. multipapillatum s. l. en
mamíferos y por lo tanto en el hombre, está influenciada por la temperatura en la
que se desarrollan sus larvas, como sucede en el nemátodo Echinocephalus
sinensis, que presenta mayor capacidad infectiva cuando se desarrolla en
temperaturas de 28 y 33 ºC.
Es necesario realizar un estudio que demuestre si existen infecciones por
C. multipapillatum s. l. o reacciones alérgicas a sus antígenos, en la gente que
consume pescado de la localidad.
También es necesario realizar pruebas de reacción cruzada entre los
antígenos de C. multipapillatum s. l. y los encontrados en otros nemátodos y
helmintos parásitos de peces, ya que existen estudios que han demostrado que
la presencia de nemátodos parásitos, puede provocar, en el hospedero, la
producción de anticuerpos, que reconocen antígenos de otros parásitos.
Debido a que algunos de los hospederos naturales de parásitos,
reconocen una menor cantidad de antígenos que los que son capaces de
Page 54
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
46
reconocer hospederos accidentales o experimentales, es posible determinar la
producción de anticuerpos contra el parásito en sus hospederos intermediarios
naturales peces, realizando pruebas de inmunoreconocimiento, para entender
mejor los procesos de infección de la especie y la respuesta de sus hospederos.
Además de pruebas que demuestren si C. multipapillatum s. l. presenta
antígenos resistentes a temperaturas altas, como se ha demostrado en otras
especies de anisákidos (Anisakis simplex).
LITERATURA CITADA
Aho, J. M. y A. O. Bush. 1993 Community richness in parasites of some
freshwater fishes from North America. In Ricklefs, R. E. and Schluter, D.
(ed.) Species diversity in ecological communities: historical and
geographical perspectives. University of Chicago Press. 185 - 190 pp.
Al-Zubaidy, A. B. 2009. Prevalence and Densities of Contracaecum sp. Larvae in
Liza abu (Heckel, 1843) from Different Iraqi Water Bodies. Marine Science.
20: 3 -17.
Anderson, R. 2000. Nematode parasites of vertebrates. Their Development and
transmission. 2nd Edition. CABI. UK. 650 pp.
Angot, V. y P. Brasseur. 1995. Anisakid larvae and their incidence on fish quality.
Revue de Medecine Veterinaire. 146 (12): 791 - 804.
Audicana, M. T., I. Ansotegui, L. F. de Corres y M. W. Kennedy. 2002. Anisakis
simplex: dangerous - dead and alive?. Trends in Parasitology. 18 (1): 20 -5.
Page 55
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
47
Balart, E. F., J. L. Castro-Aguirre, D. Aurioles-Gamboa, F. García-Rodríguez y C.
Villavicencio-Garayzar. 1995. Adiciones a la ictiofauna de la Bahía de La
Paz, Baja Califoria Sur, México. Hidrobiológica. 5 (1-2): 79-85.
Bergmann, G. T. y P. J. Motta. 2004. Infection by Anisakid Nematodes
Contracaecum spp. in the Mayan Cichlid Fish “Cichlasoma (Nandopsis)”
urophthalmus (Günther 1862). Journal of Parasitology. 90 (2): 405 - 407.
Berland, B. 1989. Identification of larval nematodes from fish. Nematode problems
in North Atlantic fish. Reporte de taller en Kiel 3-4 April 1989. Council
Meeting, Session F, Paper No 6: 16-22.
Bouree, P., A. Paugam y Petithory. 1997. Anisakidosis: report of 25 cases and
review of the literature. A collection of publications on nematodes occurring
in cod fish. Comparative Immunology, Microbiology and Infectious
Diseases. 18 (2): 75 - 84.
Bradford, M. 1976. A Rapid and Sensitive Method for the Quantitation of
Microgram Quantities of Protein Utilizing the Principle of Protein-Dye
Binding" Analytical Biochememistry. 72: 248 - 254.
Bush, A. O., K. D. Lafferty, J. M. Lotz y A. W. Shostak. 1997. Parasitology meets
ecology on its own term: Magolis et al. Revisited. Journal of Parasitology.
83: 575 - 583.
Calderón-Tinoco J. 1984. Fisiología III, Inmunología. Compañía Editorial
Continental, S.A. de C.V. México. 73 pp.
Camacho, M. 1996. Estudios de contaminación bacteriológica del cuerpo de agua
de la Ensenada de La Paz. Tesis de Licenciatura. Biología Marina.
UABCS, La Paz, B.C.S. México. 82 pp.
Page 56
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
48
Castillo Sánchez, E. y C. Pérez Ponce de León. 1998. Helminth parasites of
Paralichthys californicus (Osteichthyes: Paralichthydae) in Estero de
Punta Banda, Todos Santos Bay and San Quintin Bay, Baja California,
México. Ciencias Marinas. 24 (4): 443 - 462.
Contreras, F. 1985. Las Lagunas Costeras Mexicanas. Centro de Ecodesarrollo y
Secretaria de Pesca. México. 253 pp.
Coscia, M. y U. Oreste. 2001. Presence of antibodies specific for proteins of
Contracaecum osculatum (Rudolphi, 1908) in plasma of several Antarctic
teleosts. Fish & Shellfish Immunology. 8 (4): 295 - 302.
Cruz-Orozco, R. C. Martínez-Noriega y A. Mendoza-Maravillas. 1996. Batimetría
y sedimentos de la Bahía de La Paz, B.C.S. Oceánides. 11(1): 21-27.
Cuellar, C., M. Perteguer y M. Rodero. 2001. Presence of IL-4-Like molecules in
larval excretory-secretory products and crude extracts from Anisakis
simplex. Scandinavian Journal of Inmunology. 53 (5): 483-488.
Deardorff, T. L., y R. M. Overstreet. 1980. Contracaecum multipapillatum (=C.
robustum) from fishes and birds in the northern Gulf of Mexico. Journal of
Parasitology. 66: 853 - 856.
Del Pozo, V., I. Arrieta, T. Tunon, I. Cortegano, B. Gomez, B. Cardaba, S.
Gallardo, M. Rojo, G. Renedo, P. Palomino, A. Tabar y C. Lahoz. 1999.
Immunopathogenesis of human gastrointestinal infection by Anisakis
simplex. The Journal of Allergy and Clinical Immunology. 104 (3): 637 - 43.
Escalante H., R. Liñan, E. Díaz, K. Davelois y O. Huamanchay. 2005. Antígenos
de larvas pulmonares de Ascaris suum reconocidos por anticuerpos
Page 57
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
49
producidos por Oryctolagus cuniculus. Parasitología Latinoamericana. 60:
132 - 137.
Espinoza J. y H. Rodríguez. 1987. Seasonal phenology and reciprocal
transplantation of Sargassum sincola Setchell Gardner in the southern Gulf
of California. Journal Experimental Marine Biology Ecology. 110: 183 - 195.
Fischer W., F. Krupp, W. Schneider, C. Sommer, K. E. Carpenter y V. H. Niem.
Guía para identificación de especies para los fines de pesca. Organización
de las Naciones Unidas para la Agricultura y la Alimentación. Roma.
605 pp.
Flores, H. 1995. Parásitos de las “Cabrillas” del género Mycteroperca (Gill, 1863)
en el sur de la Península de Baja California, México. Tésis de Licenciatura.
Biología Marina, UABCS, La Paz, B.C.S. 147 pp.
Gómez-Sáenz, J. T., M. J. Gérez, M. R. Zángroniz, E. Muro, J. J. González y M.
J. García. 1999. Reacciones de hipersensibilidad y manifestaciones
digestivas producidas por la ingestión de pescado parasitado por Anisakis
simplex. Semergen. 25: 792 - 797.
Gómez del Prado Rosas, M.C., L. Iglesias, F. J. Adroher y A. Valero. 2008.
Contracaecum sp. (Nematoda: Anisakidae) parásito de lisas ¿Riesgo
potencial en la salud pública de La Paz, B.C.S? Memorias del 1er.
Congreso Peruano de Helmintología e Invertebrados Afines – Encuentro
Internacional “New approaches about Neotropical Helminthology”. 30 de
octubre - 01 de noviembre de 2008. Lima, Perú. (Resumen)
Page 58
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
50
Huizinga, H. W. 1967. The Life Cycle of Contracaecum multipapillatum (von
Drasche, 1882) Lucker, 1941 (Nematoda: Heterochelidae). The Journal of
Parasitology. 53 (2): 368 - 375.
Iannacone, J y L. Alvariño. 2009. Metazoos parásitos de Mugil cephalus
Linnaeus, 1758 (Mugilidae: Perciformes) procedentes del Terminal
Pesquero de Chorrillos, Lima, Perú. Neotropical Helminthology.
3 (1): 15 - 28.
Isishukura, H. 1989. General survey of Anisakis and anisakiasis in Japan. In
Gastric anisakiasis in Japan. Ishikura H y Namiki M. (ed.). Springer Verlag.
Tokio. 3-11 pp.
Juárez, A. 1986. Helmintos de la lisa Mugil cephalus Linneus, en Topolobampo,
Sinaloa SEDEMAR. Dirección General de Oceanografía, Topolobampo,
Sinaloa. 56 pp.
Juárez-Arroyo, J. y G. Salgado-Maldonado. 1989. Helminths of the "lisa" Mugil
cephalus at Topolobampo, Sinaloa, México. Anales del Instituto de
Biología, Universidad Nacional Autónoma de México. Serie Zoología.
México. 60 (3): 279-298.
Kato, K., N. Kagei y Y. Hayashi, Ando. 1992. Parasitological and epidemiological
survey of anisakid larvae from sardines, Engraulis japonica, caught in the
sea near Kamogawa City, Chiba Prefecture, Japan, where human
anisakiasis prevailed. Japanese Journal of Parasitology. 41 (5): 425 - 430.
Kim, L. S., Y. H. Lee, S. Kim, H. R. Park y S. Y. Cho. 1991. A case of anisakis
causing intestinal obstruction. Korean Journal of Parasitology. 29 (1): 93 -
96.
Page 59
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
51
Ko, R. C. 1976. Experimental infection of mammals with larval Echinocephalus
sinensis (Nematoda: Gnathostomatidae) from oysters (Crassostrea gigas).
Canadian Journal of Zoology. 54: 597 - 609.
Ko, R. C. 1977. Effects of temperature acclimation on infection of Echinocephalus
sinensis (Nematoda: Gnathostomatidae) from oysters to kittens. Canadian
Journal of Zoology. 55: 1129 - 1132.
Labriola J, D. y M. Suriano. 1996. Parasitic nematodes of birds from De Monte
Pond, Buenos Aires, Argentina. Boletín Chileno de Parasitología. 51:59 -
65.
López-Arellano M. E., E. Liébano-Hernández y P. Mendoza de Gives. 2005.
Haemonchus contortus L4: Desarrollo in vitro y análisis de antígenos con
posible potencial inmunoprofiláctico. Memorias del Congreso Nacional de
Buiatria XXIX, 2005. (Resumen)
López Penas, D., L. M. Ramirez Ortiz, R. del Rosal Palomeque, F. López Rubio,
R. Fernandez-Crehuet Navajas y G. Mino Fugarolas. 2000. Anisakiasis in
Spain: an increasing disease. Review. Gastroenterología y Hepatología. 23
(6): 307-11.
López, A. M., R. C. Duarte, A. Reyes-Salinas y J. E. Valdés Holguín. 2001.
Cambio Estacional de Clorofila en la Bahía de La Paz, B.C.S., México.
Hidrobiológica. 11 (1): 45 - 52.
Lorenzo, S., F. Romaris, R. Iglesias, M.T. Audícana, J. Alonso, J. Leiro y F.M.
Ubeira. 2000. O-glycans as a source of cross-reactivity in determinations of
human serum antibodies to Anisakis in determinations of human serum
Page 60
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
52
antibodies to Anisakis simplex antigens. Clinical & Experimental Allergy. 30
(4): 551 - 559.
Lu Junyi, Wu Jinying, Yang Dawei, Chen Zhisheng y Zeng Hua. 2001. Analysis of
helminth population and community composition parasitic in striped mullet
(Mugil cephalus). Acta zoologica sinica/Dongwu Xuebao. Beijing. 47 (6):
609 - 615.
MacDonald A. S., M. Araujo y E. J. Pearce. 2002. Immunology of parasitic
helminth infections. American Society for Microbiology. 70 (2): 427 - 233.
Martínez-Díaz S. y M. Hernández Rivas. 2010. Sistema para el monitoreo de la
calidad del agua. Ciencia, tecnología e innovación para el desarrollo de
México. Nolasco-Soria H. (ed.) Año 2, No. 52.
Matsuoka, H., T. Nakama, H. Kisanuki, H. Uno, N. Tachibana, H. Tsubouchi y Y.
Horii, Nawa. 1994. A case report of serologically diagnosed pulmonary
Anisakiasis with pleural effusion and multiple lesions. American Journal of
Tropical Medicine and Hygiene. 51 (6): 819 - 822.
Mattiucci S., J. Olivero, M. Paoletti, B. Arroyo y G. Nascetti. 2006. Genetic
evidence for new species of Contracaecum (Nematoda, Anisakidae)
parasites of the brown pelican, Pelecanus occidentalis, from Columbia:
genetic relationships between congeners and larval identification. 11th
International Congress of Parasitological Associations, 6th –11th August,
Glasgow Scotland. (Resumen)
Mattiucci S., M. Paoletti, A. Consuegra y G. Nascetti. 2010. Contracaecum
gibsoni n. sp. and C. overstreeti n. sp. (Nematoda: Anisakidae) from the
Page 61
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
53
Dalmatian pelican Pelecanus crispus (L.) in Greek waters: genetic and
morphological evidence. Systematic Parasitology. 75: 207 - 224.
Mattiucci, S. y G. Nascetti. 2008. Advances and Trends in the Molecular
Systematics of Anisakid Nematodes, with implications for their Evolutionary
Ecology and Host-Parasite Co-evolutionary Processes. En Advances in
Parasitology, Rollinson, D. y S. I. Hay (Ed). Elsevier. 301 pp.
Mbahinzireki, G. B. 1980. Observation on some common parasites of Bagrus
docmac Forskahl (Pisces: Siluroidea) of Lake Victoria. Hydrobiology. 75:
273 - 280.
Meléndez, S. y M. Rosas. 1995. Algunos aspectos ecológicos de los helmintos
que afectan a las especies de peces endémicas del lago de Pátzcuaro
Michoacán, México. Tésis, Facultad de Ciencias, UNAM, México. 83 pp.
Mendoza, C., L. García y Pérez-Ponce de León. 1996. Helmintos de la
“acúmara” Algansea latirostris en el lago de Pátzcuaro Michoacán, México.
Anales del Instituto de Biología, UNAM, México. Serie Zoológica. 67 (1):
71-88.
Mhaisen, F. T., N. K. Al-Salim y N. R. Khamees. 1988. Occurrence of parasites of
the freshwater mugilid fish Liza abu (Heckel) from Basrah, Southern Iraq.
Journal of Fish Biology. 32: 525 - 532.
Moravec, F., C. Vivas-Rodríguez, T. Scholz, J. Vargas-Vázquez, E. Mendoza-
Franco, J.J. Schmitter-Soto y D. González-Solís. 1995. Nematodes
parasitic in fishes of cenotes (=sinkholes) of the Peninsula of Yucatan,
Mexico. Part 1. Adults. Folia Parasitologica. 42: 115-129.
Page 62
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
54
Moreno, I., M. Caballero, F. Gomez y E. Ortega and E. Alonso. 2000. Isolation
and characterization of a major allergen from the fish parasite Anisakis
simplex. Journal of Allergy and Clinical Immunology. 106 (1): 177-82.
Moreno-Ancillo, A., M. Caballero, R. Cabañas, J. Contreras, J. Martin-Barroso, P.
Barranco y P. Lopez-Serrano. 1997. Allergic reactions to Anisakis simplex
parasitizing seafood. Annals of Allergy, Asthma & Immunology. 79 (3): 246
- 250.
Muhlia-Melo, A. y A. Muhlia-Almazan. 1995. Growth and Survival of the Lisa fish,
Mugil curema, Related to Salinity and Temperature in Experimental
Conditions in La Paz, Baja California Sur, Mexico. Aquaculture a95.
(Resumen)
Nadler S. A., S. D'Amelio, M. D. Daileyt, L. Paggi, S. Siu y J. A. Sakanari. 2005.
Molecular phylogenetics and diagnosis of Anisakis, Pseudoterranova, and
Contracaecum from Northern Pacific marine mammals. Journal of
Parasitology. 91 (6): 1413 - 1429.
Noelle, H. 1998. (Clinical aspects of the Anisakiasis of human beings).
(Proceedings of the food science advisory board for the German fisheries.
31. Annual meeting), verhandlungen des ernaehrungs-wissenschaftlichen
beirats der deutschen fischwirtschaft. 31 (13): 77-86.
Noh, J. H., B. J. Kim, S. M. Kim, M. S. Ock, M. I. Park y J. Y. Coo. 2003. A case of
acute gastric anisakiasis provoking severe clinical problems by multiple
infection. Korean Journal of Parasitology. 41: 97 - 100.
Obeso-Nieblas M. y A. R. Jiménez-Illescas. 1989. Propagación de la
constituyente M2 de la marea en La Bahía de La Paz, Baja California Sur,
Page 63
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
55
México, mediante un modelo bidimensional hidrodinámico numérico.
Investigaciones Marinas, CICIMAR. 4: 241 - 256.
Obeso-Nieblas M., B. Shirasago-Germán, J. Gaviño-Rodríguez, E. Perez-
Lezama, H. Obeso-Huerta y A. Jiménez-Illescas. Variabilidad hidrográfica
en Bahía de La Paz, Golfo de California, México (1995-2005). Revista de
Biología Marina y Oceanografía. 43(3): 559 - 567.
Olivero-Verbel J., R. Baldiris-Ávila, J. Güette-Fernaández, A. Benavides-Álvarez,
J. Mercado-Camargo y B. Arroyo-Salgado. 2006. Contracaecum sp.
infection in Hoplias malabaricus (moncholo) from rivers and marshes of
Colombia. Veretinary Parasitology. 140: 90 - 97.
Perec, A. y A. Okulewicz. 2006. The presence of Syphacia obvelata in laboratory
mice (BALB/c) – parasite antigens in immune response. Helminthologia.
43 (4): 203 - 207.
Pérez, S. y A. Ruiz-Luna. 1985. Los animales comestibles de importancia
comercial en aguas mexicanas. CECSA. México. 213 pp.
Pérez-Pérez. J., E. Fernández-Caldas, F. Marañón, J. Sastre, M. L. Bernal, J.
Rodríguez y C. A. Bedate. 2000. Molecular cloning of paramyosin, a new
allergen of Anisakis simplex. International Archives of Allergy and
Immunology. 123 (2): 120 - 129
Pérez-Ponce de León, D. García-Prieto, Osorio-Sarabia y V. León-Regagnon.
1996. Listados Faunísticos de México VI. Helmintos parásitos de peces de
aguas continentales de México. Instituto de Biología, UNAM. 100 pp.
Pérez-Ponce de León, G. 1992. Sistemática del género Pogthodiplostomum,
Dubois, 1936 y algunos aspectos epizotiológicos de la Postodiplostomiasis
Page 64
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
56
en el lago de Pátzcuaro, Michoacán, México. Tesis doctoral, Facultad de
Ciencias, UNAM, México. 181 pp.
Plomozo-Lugo T. 2010. Propuesta para la regionalización de la pesca ribereña
en el Golfo de California. Tesis de Licenciatura. Biología Marina, UABCS,
La Paz, B.C.S. 42 pp.
Ranzani-Paiva, M. y M. Tavares-Días. 2002. Erythrogram, hepatosomatic,
splenosomatic and viscerosomatic relation in mullet, Mugil platanus
Guenther (Osteichthyes, Mugilidae) parasitized. Revista Brasileira de
Zoología. 19 (3): 807 - 818.
Rivas, L. 1980. Synopsis of knowledge on the taxonomy, biology, distribution and
fishery of the Gulf of Mexico mullets (Pisces: Mugilidae). Proceedings of a
workshop for potential fishery resources of the northern gulf of México.
New Orleans, Louisiana. 34 - 53 pp.
Roberts, R. J. 1981. Patología de los peces. Mundi-Prensa. Madrid. 333 pp.
Robles Gil-Mestre S. 1998. El clima de la ciudad de La Paz. Tesis de Maestría.
UNAM. 233pp.
Rodero, M., T. Jiménez, T. Chivato, R. Laguna y C. Cuellar. 2001. Purification of
Anisakis simplex antigen by chromatography. Parasitology Research. 87:
736 - 740.
Rodríguez Romero, J., L. Abitia Cárdenas, F. Galván Magaña y H. Chávez
Ramos. 1994. Composition, abundance and specific richness of fishes from
Concepcion Bay, Baja California Sur, México. Ciencias marinas. 20 (3):
321 - 350.
Page 65
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
57
Rohde, K. 1993. Ecology of marine parasites: An Introduction to Marine
Parasitology, 2a Ed. CAB Interational. 298 pp
Rueda, S. 1995. Patrón de precipitación en la península de Baja California,
México (1894-1981). Investigaciones Marinas, CICIMAR. 10: 43 - 50.
Sakamari, J. A. 1990. Anisakis from the platter to the microfuge. Parasitology
Today. 6 (10): 323 - 327.
Sakanari, J. A. y J. H. Mckerrow. 1989. Anisakiasis. Clinical Microbiology
Reviews. 2: 278 - 284.
Salgado, M., R. Pineda, V. Vidal y C. Kennedy. 1997. A checklist of Metazoa
parasites of cichlid fish from Mexico. Journal of Helminthological Society of
Washington. 64: 195 - 207.
Salgado-Maldonado, G. y N. Barquín-Álvarez. 1978. Floridosentis elongatus
Ward, 1953 and Contracaecum sp., parasitic on Mugil cephalus Linnaeus,
1758. Anales del Instituto de Biología, Universidad Nacional Autónoma de
México. Zoología. 49 (1): 71 - 82.
Sánchez-Rueda, P. 2002. Stomach content analysis of Mugil cephalus and Mugil
curema (Mugiliformes: Mugilidae) with emphasis on diatoms in the
Tamiahua Lagoon, México. Revista de Biología Tropical. 50 (1): 245 - 252.
Shamsi S., R. Gasser, I. Beveridge y A. A. Shabani. 2008. Contracaecum
pyripapillatum n. sp. (Nematoda: Anisakidae) and a description of C.
multipapillatum (von Drasche, 1882) from the Australian pelican, Pelecanus
conspicillatus. Parasitology Research. 103: 1031 - 1039.
Page 66
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
58
Shamsi S., R. Norman, R. Gasser y I. Beveridge. 2009. Redescription and genetic
characterization of selected Contracaecum spp. (Nematoda: Anisakidae)
from various hosts in Australia. Parasitology Research. 104: 1507 - 1525.
Skirnisson, K. 2006. Pseudoterranova decipiens (Nematoda: anisakidae) larval
reported from humans in Iceland after consumption of insufficiently cooked
fish. Laeknabladid. 92: 21 - 25.
Stoskopf, M. 1993. Fish Medicine. Saunders Company. U.S.A. 882 pp.
Stuardo, J. y A. Martinez. 1975. (General results of a biological and fishery survey
of the coastal lagoons of the State of Guerrero, Mexico). Acta Politécnica.
16 (72): 99 - 115.
Sugimachi, E., K. Inocuchi, T. Iowa, T. Fujino y Y. Ishii. 1985. Acute gastric
anisakiasis: Análisis of 128 cases. Journal of American Medicine. 253 (7):
1012 - 1013.
Szalai, H. y P. E. Dick. 1990. Proteocephalus ambloplitis and Contracaecum sp.
From Largemouth Bass (Micropterus salmoides) stocked into Boundary
reservoir, Saskatchewan, Journal of Parasitology. 76 (4): 598 - 601.
Tizard, I. R. 2002. Inmunología Veterinaria. 6ª Ed. McGraw-Hill Interamericana.
México. 517 pp.
Valles Ríos, M., G. Ruiz Campos y L. Galaviz Silva. 2000. Prevalence and
parasitic intensity in Mugil cephalus (Pisces: Mugilidae), from the
ColoradoRiver, Baja California, México. Revista de Biología Tropical. 48 (2-
3): 495 - 501.
Valles Vega, I. y M. C. Gómez del Prado-Rosas 2010. Cambios en la
organización anatómica de la larva L3 de Contracaceum sp. (Nematoda:
Page 67
Estudio de la estructura antigénica de Contracaecum multipapillatum s. l.
59
Anisakidae) por el empleo de distintos métodos culinarios en La Paz,
B.C.S., México. Memorias del Segundo Congreso Internacional de
Parasitología Neotropical "El Rol de la Parasitología Neotropical en la
Salud Global". 9 - 13 de Noviembre de 2010. Lima, Perú. (Resumen)
Valles-Ríos, M. E., G. Ruiz-Campos y L. Galaviz Silva. 2000. Prevalencia e
intensidad parasitaria en Mugil cephalus (Pisces: Mugilidae), del Río
Colorado, Baja California, México. Revista de Biología Tropical. 48 (2-3):
495 - 501.
Vidal-Martinez, V. M., D. Osorio-Sarabia y R. M. Overstreet. 1994. Experimental
infection of Contracaecum multipapillatum (Nematoda: Anisakinae) from
Mexico in the domestic cat. The Journal of Parasitology. 80 (4): 576 - 579.
Villaseñor-Casales, A. 1979. Distribución vertical de temperatura, salinidad y
oxigeno disuelto en La Bahía de La Paz, Baja California Sur, durante la
primavera de 1976. California Cooperative Oceanic Fisheries Investigation
Reports. 20: 146 - 149.
Warburton, K. 1979. Growth and production of some important species of fish in
a Mexican coastal lagoon system. Journal of Fish Biology. 14 (5): 449 -
464.
Vidal-Martinez, V. M., D. Pech, B. Sures, S. T. Purucker y R. Poulin. 2009. Can
parasites really reveal enviromental impact?. Trends in Parasitology. 20 (4):
170 - 177.