Maggi, Federico Javier Factores ambientales que afectan la germinación de Dipsacus fullonum L; evaluación de tempe- ratura, stress salino y osmótico Trabajo Final de Ingeniería en Producción Agropecuaria Facultad de Ciencias Agrarias Este documento está disponible en la Biblioteca Digital de la Universidad Católica Argentina, repositorio institucional desarrollado por la Biblioteca Central “San Benito Abad”. Su objetivo es difundir y preservar la producción intelectual de la Institución. La Biblioteca posee la autorización del autor para su divulgación en línea. Cómo citar el documento: Maggi, F. J. 2016. Factores ambientales que afectan la germinación de Dipsacus fullonum L; evaluación de temperatura, stress salino y osmótico [en línea]. Trabajo Final de Ingeniería en Producción Agropecuaria. Facultad de Ciencias Agrarias. Universidad Católica Argentina. Disponible en: http://bibliotecadigital.uca.edu.ar/repositorio/tesis/factores-ambientales-afectan-germinacion.pdf [Fecha de consulta:.........]
Transcript
Maggi, Federico Javier
Factores ambientales que afectan la germinación de Dipsacus fullonum L; evaluación de tempe-ratura, stress salino y osmótico
Trabajo Final de Ingeniería en Producción Agropecuaria Facultad de Ciencias Agrarias
Este documento está disponible en la Biblioteca Digital de la Universidad Católica Argentina, repositorio institucional desarrollado por la Biblioteca Central “San Benito Abad”. Su objetivo es difundir y preservar la producción intelectual de la Institución.La Biblioteca posee la autorización del autor para su divulgación en línea.
Cómo citar el documento:
Maggi, F. J. 2016. Factores ambientales que afectan la germinación de Dipsacus fullonum L; evaluación de temperatura, stress salino y osmótico [en línea]. Trabajo Final de Ingeniería en Producción Agropecuaria. Facultad de Ciencias Agrarias. Universidad Católica Argentina. Disponible en: http://bibliotecadigital.uca.edu.ar/repositorio/tesis/factores-ambientales-afectan-germinacion.pdf [Fecha de consulta:.........]
PONTIFICIA UNIVERSIDAD CATÓLICA ARGENTINA
Facultad de Ciencias Agrarias
Ingeniería en Producción Agropecuaria
“Factores ambientales que afectan la germinación de Dipsacus fullonum L;
evaluación de temperatura, stress salino y osmótico”
Trabajo Final de Graduación para optar por el título de:
Ingeniero en Producción Agropecuaria
Autor: Maggi, Federico Javier
Profesor tutor: Huarte, Hector Roberto
Modalidad: Experimental
Fecha: Febrero 2016
2
10 de febrero del 2016
Sr. Director de la carrera de IPA Presente.- De mi consideración:
Tengo el agrado de dirigirme a Ud., adjuntando el Trabajo Final “Factores ambientales que afectan la germinación de Dipsacus fullonum L; evaluación de temperatura, stress salino y osmótico” (dos copias), que ha sido desarrollado por el alumno Federico Javier Maggi, Nro. de Registro 05-100034-9, en cumplimiento de las disposiciones vigentes.
Dicho trabajo ha sido realizado bajo mi dirección y, habiendo evaluado el mismo, lo considero aprobado con los siguientes comentarios.
Por lo expuesto anteriormente, avalo su presentación ante el Comité Evaluador correspondiente.
Sin otro particular lo saludo atte.
Firma
Aclaración
Cargo Docente IPA
3
Agradecimientos
A Hector Roberto Huarte, que en su rol de tutor hizo todo lo necesario para terminar este proyecto en tiempo y forma
A Margarita Solivella por su ayuda en el laboratorio durante los ensayos
A mis abuelos, padres y hermanos que son mi guía y sustento
A Eduardo y Susana por su apoyo e interés en todos estos años
A Carlos y Zulma que sin ellos no hubiese llegado a esta instancia
A mis amigos que siempre me apoyaron y ayudaron
4
Índice
Resumen página 5
Introducción página 6
Objetivos página 10
Materiales y Métodos página 11
Resultados y Discusión página 13
Conclusión página 20
Bibliografía página 21
5
Resumen
Dipsacus fullonum es una especie conocida vulgarmente como carda, carda silvestre
o cardencha, perteneciente a la familia Dipsacaceae. Es una maleza muy invasora que
crece en sitios abiertos y resiste condiciones de sequía, inundación y salinidad.
Conocer la tolerancia de esta especie a estos factores permitiría conocer su potencial
área de infestación. Al presente esta información es escasa en la Argentina. Por otro
lado, tampoco se dispone de información acerca del valor de sus temperaturas
cardinales ni de la tolerancia al estrés salino u osmótico. Por ello, dentro de este
marco teórico se evaluaron: i) el potencial agua base de germinación (ψb (50))
aplicando el modelo del hidrotiempo y ii) las temperaturas cardinales y iii) el tiempo
térmico a germinación (TT). Para la obtención de estos parámetros se condujeron
diferentes ensayos de germinación a las siguientes temperaturas: 5°C; 10°C; 15°C;
20°C; 25°C; 30°C y 35°C. Se calculó el porcentaje de semillas germinadas y el
tiempo medio a germinación para todos los tratamientos térmicos. Fue observado que
a medida que la temperatura aumentó, disminuía el tiempo medio a germinación
(T50) y sólo en dos temperaturas estudiadas el porcentaje final no llegó al 100% (i.e.
5°C y 30°C) y a 35°C no se observó germinación. La temperatura base fue de 4,94ºC,
la óptima fue 25,46ºC y la crítica fue 38,90ºC. El tiempo térmico para la etapa de
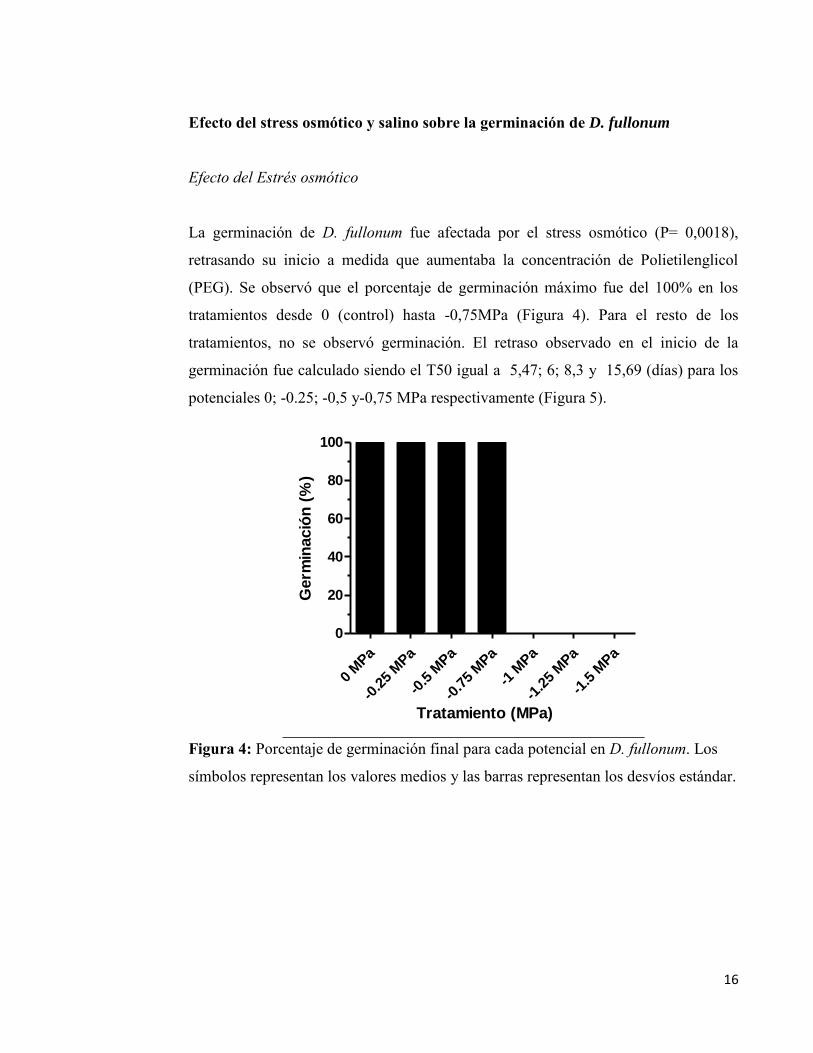
germinación fue de 39,06ºCd. En los ensayos de stress osmótico y salino, se observó
que a medida que las semillas fueron incubadas a potenciales más negativos el
porcentaje de germinación final no varió excepto en los potenciales muy negativos (-
1 MPa en NaCl y -0,75MPa en PEG). En cambio, se observó un retraso en el inicio
de la germinación. Estos resultados proponen que en coincidencia con sus
características morfológicas, su hábito de crecimiento, su fácil dispersión y el amplio
rango de condiciones ambientales donde la germinación alcanza elevados porcentajes
permiten proponer que D. fullonum sea reportada en nuevas zonas.
6
Introducción
Dipsacus fullonum es una especie conocida vulgarmente como carda, carda silvestre
o cardencha, perteneciente a la familia Dipsacaceae. Es una maleza muy invasora que
crece en sitios abiertos, resiste condiciones de sequía e inundación (Dimitri, 1985).
Dipsacus es nativa de Europa y se la encuentra también en los Estados Unidos y
Canadá (Glass, 2007). Desde Ontario, se esparció 650 km hasta la costa este de
Estados Unidos en solamente 13 años (Bentivegna, 2008), donde está presente en 43
de las 48 provincias de dicho país (Glass, 2007). En la Argentina está reportada en las
provincias de Buenos Aires, Córdoba, Neuquén, Rio Negro, Jujuy (Burkart, 1957). Es
muy agresiva e invade rápidamente una pradera con el agravante que carece de
enemigos naturales (Glass, 2007). Además, es hospedante secundario de plagas que
atacan cultivos, como el Virus del Mosaico del Girasol (Giolitti et al, 2009). Su
establecimiento y posterior crecimiento es alto debido a sus características
morfológicas, como una raíz pivotante profunda y hojas muy espinosas que evitan la
defoliación por el ganado. Las rosetas establecidas cubren el suelo rápidamente,
reduciendo el espacio y la luz para otras especies (Bentivegna, 2008). Es una especie
bianual que se reproduce por semillas (Busso, 2013) y no se registró reproducción
vegetativa (Sforza, 2004). Su hábito de crecimiento es en roseta con hojas
lanceoladas con espinas en la nervadura central, en la cara inferior que puede alcanzar
los 60 cm de diámetro en el primer año. En la siguiente temporada de crecimiento,
produce un vástago o tallo floral, que puede medir entre 0,5 a 2,5m de altura y en su
extremo posee un capitulo cilíndrico de 2,5 a 10 cm de largo. Las hojas anteriores a la
inflorescencia son opuestas, permitiendo a la planta contener el agua de lluvia
(Werner, 1975). Además del tallo principal, posee ramas secundarias que también
producen capítulos (Topham, 1968). Rodeando el capítulo, se encuentran las brácteas
espinosas en la nervadura central que son más largas que el capítulo y se curvan. Las
flores poseen el cáliz reducido y la corola lila que está formada por cuatro pétalos
fusionados. Posee ovario ínfero que produce un sólo fruto, un aqueño. Dicho aqueño
posee 4 o 5 milímetros de largo, y se encuentra adherido a un papus de color marrón.
7
Las aquenios son dispersados en su mayoría cerca de la planta madre, pero también
son dispersadas por el agua, animales, humanos y vehículos. Por este motivo, nuevas
áreas pueden verse afectadas, formando manchones (Busso, 2013). Produce más de
3000 semillas por planta (Werner, 1975). Fue reportado para poblaciones de semillas
recolectadas en los EE. UU que esta especie tolera altos niveles de salinidad durante
la germinación (Beaton y Dudley, 2013). No obstante, carecemos de información
sobre el comportamiento frente a este factor de poblaciones argentinas.
Las relaciones hídricas que se establecen entre la semilla y el suelo condicionan el
porcentaje y velocidad de germinación (Bradford, 2002). Suelos bien provistos de
agua permiten que este proceso suceda más rápidamente. Asimismo, las
características estructurales de los suelos modifican la cantidad de agua retenida y la
facilidad a desprenderse de ella. Si bien muchas veces se utilizan como sinónimos, los
suelos salinos y alcalinos presentan caracterizas diferentes. En el primer caso, es un
suelo que contiene gran cantidad de sales solubles, suficientes para afectar el
crecimiento de las plantas. Su límite es estimado en 4 dS m-1. En cambio, los suelos
sódicos, son los que poseen bajo nivel de sales totales, pero contienen niveles de
sodio que afectan adversamente la estructura del suelo y a la vegetación presente. En
ambos casos se ve reducido el potencial agua. El potencial agua es la capacidad que
tiene el agua de realizar trabajo, es decir, su energía, por convención se lo considera
negativo (a medida que el suelo se seca, hay menos agua disponible, haciéndose más
negativo). Este potencial, se simboliza con la letra ψw (psi del alfabeto griego) y es la
sumatoria de cuatro potenciales, el potencial del soluto u osmótico (ψs), pues estos
interaccionan con el agua disminuyendo su energía, el potencial de presión (ψp) que
es la presión hidrostática a la que está sometida el agua, el potencial de gravedad (ψg),
que permite el movimiento del agua hacia abajo del perfil de suelo y el potencial
matricial o mátrico (ψm) que es la adsorción del agua por parte de los componentes
del suelo (Álvarez, 2012). Es de gran importancia conocer el valor del potencial agua,
ya que permite saber si esta se mueve o permanece estática, pues el agua entra en la
célula siguiendo un gradiente de presión (Taiz, 2006), de esta manera es
8
imprescindible conocer ciertos valores de potencial agua, que se usan como
referencia, y así estimar si una planta está con posibilidades de sufrir stress. Estos
valores son tres; potencial 0 MPa, que representa el agua libre es el estado de máxima
energía, -0,033 MPa que representa la capacidad de campo, que se la define como la
cantidad de agua retenida en el suelo, luego de drenar el agua gravitacional y
finalmente el -1,5 MPa que es el punto de marchitez permanente, se refiere al
contenido de agua donde la energía de retención es tan alta que los vegetales no la
pueden absorber (Álvarez, 2012). El estrés osmótico ocasionado por la presencia de
sodio, trae aparejado una serie de inconvenientes para la planta, ya que este compite
con el potasio en la unión con la membrana plasmática. El exceso de sodio es
retenido en la vacuola de la planta, pero cuando hay demasiado, este ocupará el
citoplasma, si el proceso continúa la célula muere por reducción del potencial agua,
es decir, el sodio atrae agua y la célula se deshidrata.
Para evaluar el comportamiento de plantas o semillas frente a una reducción del
potencial agua es común el uso de soluciones de Polietilenglicol (PEG). Esta es una
sustancia de alto peso molecular y que no puede pasar a través de la membrana
celular (Emmerich y Hardegree, 1990). El PEG no presenta toxicidad y simula la falta
de agua. Para cuantificar las respuestas al potencial osmótico de incubación,
Gummerson (1986) propuso el modelo hidrotiempo, donde el tiempo a germinación
está relacionado con el diferencial entre el potencial agua de la semilla (ψ) y el
potencial base o umbral (ψb) para la emergencia de la radícula. Se entiende por
potencial base al potencial más negativo en el que una semilla logra germinar. La tasa
de germinación de una población de semillas (GRg) está relacionada con la potencial
agua de incubación de manera linear, con una pendiente común que intercepta al eje
de las abscisas a diferentes valores. Estos puntos de intercepción se denominan
potencial agua base de germinación.
9
Como la pendiente es la misma, el hidrotiempo total (MPa-hora o MPa-día) hasta la
emergencia de la radícula será el mismo para toda la población, pero las semillas
individuales variaran en su umbral. Respondiendo a la siguiente formula:
𝜃𝐻 = (ψ − ψb(g))tg
Donde θH es la constante de hidrotiempo (tiempo requerido para que ocurra
germinación), ψ es el potencial agua de la semilla, ψb es el potencial agua base de una
población g y el tg es el tiempo a germinación del porcentaje g. Por ello, a medida que
disminuye el potencial agua, el tiempo a germinación aumentará (Bradford, 2002).
Entre más pequeña sea la diferencia entre ambos potenciales, el hidrotiempo se
acumulará más lentamente (MPa-h) llevando más tiempo para lograr germinación
(Benech-Arnold y Sánchez, 2004). Otro factor ambiental que modula la germinación
es la temperatura (Baker, 2001). La temperatura modula la velocidad del proceso de
germinación y las demás etapas del crecimiento vegetal también responden a ella
(Slafer y Rawson, 1994). La respuesta a la temperatura se representa a través del
modelo de tiempo térmico (TT), su unidad son los grados-día (ºCd) respondiendo a la
siguiente formula:
𝑇𝑇(º𝐶𝑑) = ∑(𝑇𝑚 − 𝑇𝑏)
Donde el TT es el tiempo térmico acumulado para una etapa del cultivo, Tm es la
temperatura media diaria, Tb la temperatura base (temperatura específica de cada
etapa y cultivo en la cual se detiene el desarrollo). La temperatura en la cual la
duración de la etapa es mínima (máxima tasa), se la conoce como temperatura
óptima. Si se realiza la inversa del tiempo que tardó en completarse la etapa de
germinación, se obtiene la tasa de germinación y por lo tanto se desprende que a
mayor temperatura habrá una tasa proporcionalmente mayor y por consiguiente se
requerirán menos días para completar la etapa (Satorre, 2010).
10
Objetivos
El objetivo del trabajo fue determinar el efecto de la temperatura y el potencial
osmótico y salino sobre la germinación de Dipsacus fullonum.
Objetivos particulares
i. Determinar el potencial agua base de germinación y su desviación estándar.
ii. Determinar el tiempo térmico a germinación y las temperaturas cardinales.
11
Materiales y Métodos
Las semillas utilizadas fueron recolectadas durante el mes de marzo del 2014 dentro
del predio de la Facultad de Ciencias Agrarias de la UCA (Lat. S. 34°34´, Long. O.
58°28´). Después de separar las semillas de la inflorescencia, las semillas fueron
conservadas a una temperatura de 6°C. Se realizaron tres ensayos independientes de
manera simultánea: 1) la evaluación del efecto de la temperatura sobre la
germinación, 2) la evaluación del comportamiento frente al stress salino y 3)
osmótico durante la germinación. Los datos fueron analizados mediante el análisis de
varianza (ANOVA) (InfoStat, UNRC). Las diferencias entre tratamientos fueron
detectadas utilizando el test HSD Tukey con un nivel de significancia de 0,05.
Los parámetros del modelo de hidrotiempo fueron obtenidos mediante la herramienta
Solver de Microsoft Excel® 2010. Dicha herramienta permitió lograr un mejor ajuste
entre los valores obtenidos en los experimentos con los valores simulados por el
software. El criterio utilizado para lograr el mejor ajuste fue el error cuadrático medio
entre el valor simulado y el obtenido experimentalmente.
Preparación de semillas
Para todos los ensayos, se colocaron las semillas en cajas de Petri de 9 cm de
diámetro con una hoja de papel de filtro. Cada tratamiento contó con tres repeticiones
de treinta semillas en cada una. Las semillas fueron hidratadas con 8ml de agua
destilada o la solución correspondiente. Se realizó un conteo diario de semillas
germinadas durante 28 días de incubación.
12
Efecto de la temperatura
Para determinar el efecto de la temperatura sobre la germinación se seleccionaron los
siguientes tratamientos: 5°C, 10°C, 15°C, 20°C, 25°C, 30°C y 35°C
Efecto del stress salino
Para determinar el efecto del stress salino sobre la germinación se seleccionaron los
siguientes tratamientos: 0; -0,25;-0,5;-0,75;-1;-1,25 y -1,5 MPa. Estas soluciones se
prepararon disolviendo 0,643g; 1,286g; 1,93g; 2,574g; 3,218g; 3,862g de cloruro de
sodio (NaCl, Biopack Argentina) en 200ml agua destilada respectivamente.
Efecto del stress osmótico
Para determinar el efecto del stress osmótico sobre la germinación se seleccionaron
los siguientes tratamientos: 0; -0,25; -0,5; -0,75;-1;-1,25 y -1,5 MPa. Estas soluciones
se prepararon de acuerdo con la ecuación de Michel (1983) disolviendo 26,21g;
38,37g; 47,72g; 55,61g; 62,57g; 68,85g de polietilenglicol (PEG 6000, Anedra) en
200 ml de agua destilada respectivamente.
Ambos efectos se evaluaron bajo una temperatura de 15°C constantes con un
fotoperiodo de 12h. Se realizaron reemplazos del papel de filtro y de las soluciones de
incubación para mantener el potencial agua deseado en valores estables luego de 24
horas de iniciada la incubación y de ahí en más cada 6 días.
13
Resultados y Discusión
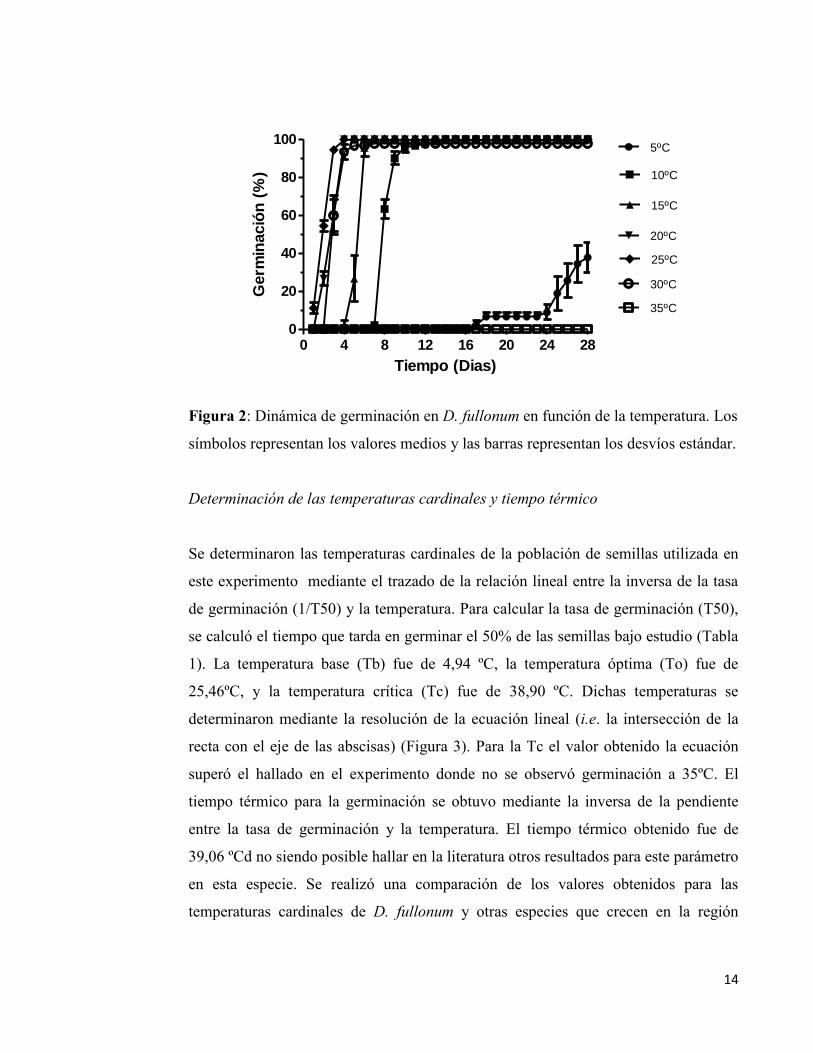
Efecto de la temperatura sobre la germinación de D. fullonum
D. fullonum germinó en un amplio rango de temperaturas, desde 5ºC hasta 30ºC. El
porcentaje de germinación alcanzó el 100% para 10ºC, 15ºC, 20ºC y 25ºC. Este
porcentaje disminuyó ligeramente (97,78 % ± 1,92%) para la temperatura de 30ºC
mientras que en el tratamiento de 5°C se redujo hasta un 37,77 % ± 13,88% (Figura
1). Por otro lado, a 35ºC no hubo germinación. Por otro lado, se observó que el
aumento progresivo de la temperatura disminuyó el tiempo a germinación (Figura 2).
En efecto el T50 calculado fue de 29,02; 7,8; 5,19;2,56;1,83 y 2,86 (días) para
5ºC,10ºC, 15ºC, 20ºC, 25ºC y 30ºC respectivamente.
5 10 15 20 25 30 35
0
20
40
60
80
100
Temperatura (ºC)
Germ
inació
n (
%)
Figura 1: Porcentaje de germinación final para cada temperatura en D. fullonum. Los
símbolos representan los valores medios y las barras representan los desvíos estándar.
14
0 4 8 12 16 20 24 280
20
40
60
80
1005ºC
10ºC
15ºC
20ºC
25ºC
30ºC
35ºC
Tiempo (Dias)
Germ
inació
n (
%)
Figura 2: Dinámica de germinación en D. fullonum en función de la temperatura. Los
símbolos representan los valores medios y las barras representan los desvíos estándar.
Determinación de las temperaturas cardinales y tiempo térmico
Se determinaron las temperaturas cardinales de la población de semillas utilizada en
este experimento mediante el trazado de la relación lineal entre la inversa de la tasa
de germinación (1/T50) y la temperatura. Para calcular la tasa de germinación (T50),
se calculó el tiempo que tarda en germinar el 50% de las semillas bajo estudio (Tabla
1). La temperatura base (Tb) fue de 4,94 ºC, la temperatura óptima (To) fue de
25,46ºC, y la temperatura crítica (Tc) fue de 38,90 ºC. Dichas temperaturas se
determinaron mediante la resolución de la ecuación lineal (i.e. la intersección de la
recta con el eje de las abscisas) (Figura 3). Para la Tc el valor obtenido la ecuación
superó el hallado en el experimento donde no se observó germinación a 35ºC. El
tiempo térmico para la germinación se obtuvo mediante la inversa de la pendiente
entre la tasa de germinación y la temperatura. El tiempo térmico obtenido fue de
39,06 ºCd no siendo posible hallar en la literatura otros resultados para este parámetro
en esta especie. Se realizó una comparación de los valores obtenidos para las
temperaturas cardinales de D. fullonum y otras especies que crecen en la región
15
observándose resultados bastante similares con Conyza bonariensis, donde se
determinaron las temperaturas de 4,2ºC; 20ºC y 35ºC como las base, óptima y crítica
respectivamente (Wu et al, 2007). En cambio, para Conyza canadensis, Amaranthus
palmeri, Digitaria sanguinalis la temperatura base calculada fue de 13ºC; 15,7ºC;
17ºC; 15,1ºC respectivamente (Steinmaus et al, 2000), en las leguminosas adaptadas a
la región, ocurre algo similar, obteniéndose para Melilotus officinalis, las
temperaturas base, optima y critica obtenidas fueron 0ºC; 18,47ºC y 34,6ºC