36
Metilación del ADN & Islas CpG

Metilación del ADN &
Islas CpG

Base bioquímica
La metilación es la adición de un grupo metilo (-CH3) a una molécula. La metilación se observa en el ADN, ARN y en proteínas.
PROCARIOTAS
EUCARIOTAS
Metil-Citosina Metil-Adenina
En eucariotas ocurre
exclusivamente en las citosinas
(en el carbono 5 - 5meC)
Frecuentemente,
metilación de la adenina en N6
Y de la citosina en C5

Metilación y desmetilación
De novo metiltrasferasa (DNMT3 ) Metila citosinas previamente no-metiladas (o hemi-metiladas)
Metiltransferasa de mantenimiento (DNMT1) Actúa principalmente sobre citosinas hemi-metiladas (metiladas en una hebra) para mantener el estado de metilación después de la replicación del DNA
Hao Wu, Yi Zhang . Reversing DNA Methylation: Mechanisms, Genomics, and Biological Functions. Cell .
Volume 156, Issue 1, Pages 45-68 (January 2014) .DOI: 10.1016/j.cell.2013.12.019

Metilación y desmetilación
Hao Wu, Yi Zhang . Reversing DNA Methylation: Mechanisms, Genomics, and Biological Functions. Cell .
Volume 156, Issue 1, Pages 45-68 (January 2014) .DOI: 10.1016/j.cell.2013.12.019
(B) Pasos de la ruta de modificación
de 5mC
C) Base excision repair of 5mC (in
plants) or 5fC/5caC (in mammals)

Desmetilación activa y pasiva
Hao Wu, Yi Zhang . Reversing DNA Methylation: Mechanisms, Genomics, and Biological Functions. Cell .
Volume 156, Issue 1, Pages 45-68 (January 2014) .DOI: 10.1016/j.cell.2013.12.019

Contexto de metilación Mamíferos: Básicamente CpG, en células no-diferenciadas también CHG y CHH
Plantas: Tanto CG como CHH y CHG
Arabidopsis thaliana Homo sapiens
IMR90: fibroblasto
H1: célula madre

Función de la metilación
Regulación de la expresión génica:
• Metilación cerca del TSS: bloquear la iniciación de la transcripción
• Metilación del cuerpo génico: Estabilización y elongación de la transcripción
Estabilidad de genoma:
• Metilación de los centrómeros
• Metilación de transposones
Desarrollo
• Desmetilación o reprogramación global de los valores de metilación en dos
fases del desarrollo
Impronta génica & inactivación del cromosoma X • Genes improntados y los ubicados en el cromosoma X desactivado suelen silenciarse
mediante metilación del DNA

Expresión génica
Relación entre nivel de expresión y probabilidad de de novo metilación
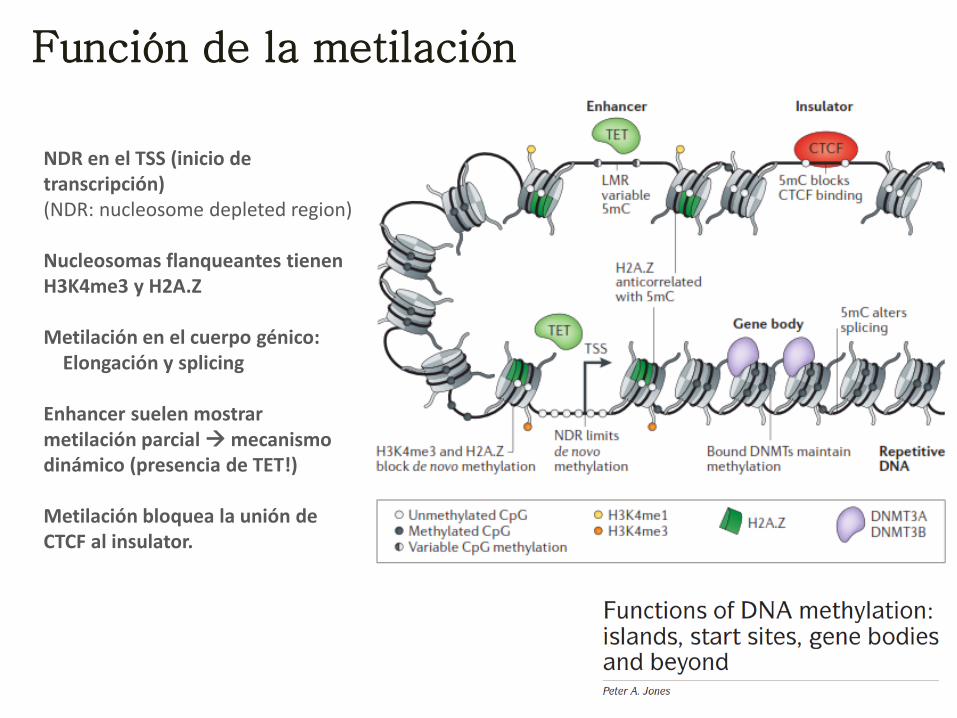
Función de la metilación
NDR en el TSS (inicio de transcripción) (NDR: nucleosome depleted region) Nucleosomas flanqueantes tienen H3K4me3 y H2A.Z Metilación en el cuerpo génico: Elongación y splicing Enhancer suelen mostrar metilación parcial mecanismo dinámico (presencia de TET!) Metilación bloquea la unión de CTCF al insulator.

Expresión génica y metilación
Que primero, el silenciamiento del gen o la metilación de la
región promotora?
1) El Promotor se metila se silencia el gen
2) El gen es silenciado el promotor se puede metilar (para estabilizar el
silenciamiento?)
1)
En cáncer, promotores con CGI son silenciados primeros y metilados después A favor de ‘silenciamiento primero’
2)
Trabajos sobre el papel de DNMT3A en la diferenciación de células
hematopoyéticos sugieren un papel mas ‘amplio’ de la metilación

Impronta génica y inactivación
Probablemente el silenciamiento de (algunos) de los genes del cromosoma X precede la metilación

Inestabilidad del genoma: cáncer
http://www.nature.com/scitable/content/dna-methylation-and-cancer-21285

Global hypomethylation in breast cancer.
Hon G C et al. Genome Res. 2012;22:246-258
Copyright © 2012 by Cold Spring Harbor Laboratory Press

Metilación y desarrollo
Figure 1. DNA methylation changes during developmental epigenetic reprogramming. Primordial germ cells (PGCs) emerge in embryos at E7.5 and, concomitant with their proliferation and migration towards the genital ridge, DNA methylation is globally erased (black line). Following sex-determination, new DNA-methylation landscapes are established in germ-cell precursors in an asymmetrical fashion in male and female embryos. In the male embryo (blue line), de novo methylation takes place before meiosis in mitotically arrested cells (G1-phase; prospermatogonia) and is completed before birth. In the female embryo (red line), primary oocytes enter meiosis and arrest in prophase-I (diplotene stage); DNA methylation is established after birth during the follicular/oocyte growth phase. At puberty, under specific endocrine triggers, fully-grown germinal vesicle (GV) oocytes resume the first meiotic division. After extrusion of the first polar body, oocytes arrest in metaphase of the second meiotic division (MII oocytes) and meiosis is completed only upon fertilisation. Following fertilisation, a new wave of DNA demethylation takes place that is distinct on the parental genomes. In the zygote, DNA methylation of the paternal genome is rapidly erased by an active mechanism (blue line). Demethylation of the maternal genome is slower (red line) and is dependent on DNA replication (passive demethylation). These post-fertilisation demethylation events do not include imprinted gDMRs (green dotted line), resulting in parental-allele-specific methylation of these elements in early embryos and consequent parental-allele-specific expression of associated imprinted genes. Concomitant with blastocyst implantation and cell-lineage determination, new methylation landscapes become established, associated with cellular differentiation.

Resumen

Publicaciones
• Deaton AM, Bird A.(2011). CpG islands and the regulation of transcription. Genes Dev. 25(10):1010-22.
• Thomson JP, Skene PJ, Selfridge J, Clouaire T, Guy J, Webb S, Kerr AR, Deaton A, Andrews R, James KD, Turner DJ, Illingworth R, Bird A. (2010).CpG islands influence chromatin structure via the CpG-binding protein Cfp1. Nature. 464(7291):1082-6.
• Suzuki, M.M. and Bird, A. (2007). DNA methylation landscapes: provocative insights from epigenomics. Nat. Rev. Genet. 9(6):465-76.
• Jones PA: Functions of DNA methylation: islands, start sites, gene bodies and beyond. Nature reviews Genetics 2012, 13(7):484-492.
• Hao Wu, Yi Zhang . Reversing DNA Methylation: Mechanisms, Genomics, and Biological Functions. Cell . Volume 156, Issue 1, Pages 45-68 (January 2014) .DOI: 10.1016/j.cell.2013.12.019

Patrón de metilación
• Los patrones de metilación (distribución a lo largo de la secuencia) no son iguales en distintos eucariotas
• La levadura (Saccharomyces cerevisiae) y C. elegans no tienen un gen homólogo al DNMT y no presentan me5C
• En fungi, solo el DNA repetido se metila • Los mayores niveles de metilación en
plantas (hasta el 50% de todas las citosinas) – metilación de contextos non-CpG en elementos transponibles
• En general encontramos un mosáico de metilación (regiones metiladas y intercaladas regiones no-metiladas
• Genomas de mamífero son predominantemente metilados (CpG) con excepción de regiones cortas llamados islas CpG
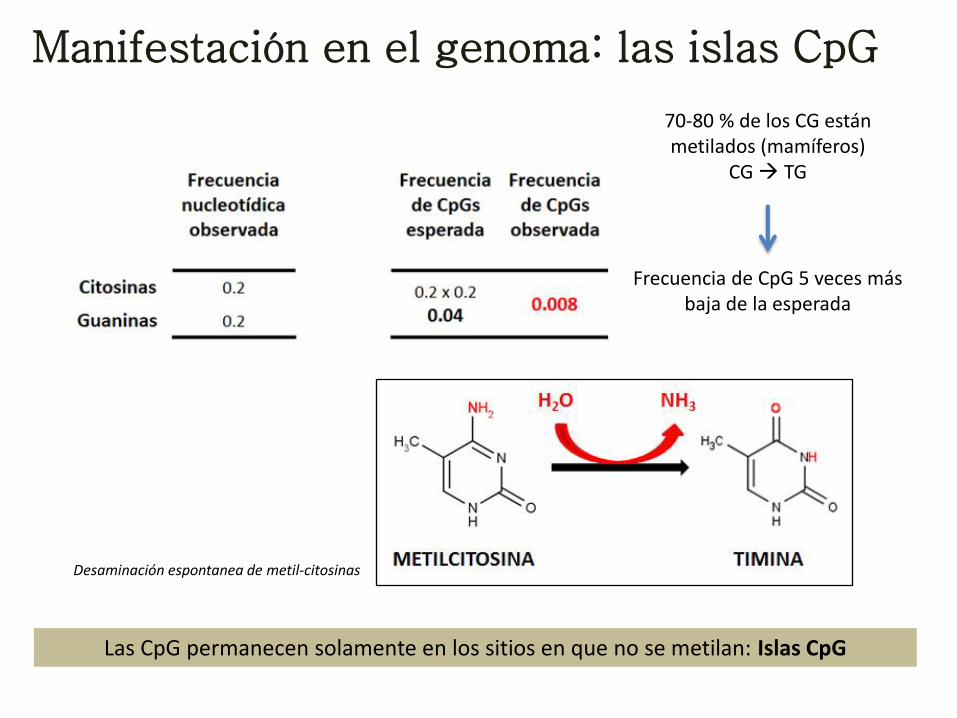
Manifestación en el genoma: las islas CpG
70-80 % de los CG están metilados (mamíferos)
CG TG
Desaminación espontanea de metil-citosinas
Frecuencia de CpG 5 veces más baja de la esperada
Las CpG permanecen solamente en los sitios en que no se metilan: Islas CpG

Propiedades de las islas CpG
1. Son ricas en G+C (ratio O/E alto) y tienen longitudes alrededor de 1kb 2. Entre el 50 y el 70% de los genes tienen una isla CpG asociada a sus promotores. 3. Casi todos los genes “housekeeping” (se expresan en todos los tejidos) tienen una isla asociada a
su promotor pero solo la mitad de los genes específicos la presentan. 4. En los promotores de los genes: Cuando se metilan dan lugar a una inhibición de la transcripción. 5. En el cuerpo génico: Cuando se metilan dan lugar a estabilización de la transcripción 6. En algunas condiciones fisiológicas o patológicas se pueden ver cambios en el estado de la
metilación: cáncer
Existen tanto métodos experimentales como computacionales para detectar islas CpG

islas CpG: ventanas deslizantes

Read the DNA sequence
Determine the distance (d) of each CpG to the next CpG downstream in the DNA
sequence:
10,5,5,3,1,8,23,34,21,12,2,5,8,6,9,...N-1
Let be dm a distance threshold If di ≤ dm Cluster seed
For example, for dm = 5:
10,5,5,3,1,8,23,34,21,12,2,5,8,6,9,...N-1
List of CpG clusters with
coordinates, length and
number of CpGs
Assign a P-value to each CpG cluster
Statistically significant cluster ≡ CpG island
CpGcluster: basado en distancias
Calculate statistical and sequence
properties: G+C content, O/E ratio, CpG
density, intra-clustering of CpGs,
overlap with Alus, PhastCons etc.
Binary sequence: CpG = 1; noCpG = 0
0001000010100000010100010100001000101010000101

¿Qué distancia usamos?

¿Cómo asignar la significación?
0 1 0 0 0 2 0 0 0 3 0 0 0 4 0 0 0
0 .0 0 0 0
0 .0 0 0 5
0 .0 0 1 0
0 .0 0 1 5
0 .0 0 2 0
0 .0 0 2 5
0 .0 0 3 0
0 .0 0 3 5
0 .0 0 4 0
N = 4 0
N = 2 0
N = 1 0
Pro
ba
bil
ity
C lu s te r le n g th (n t)
R a n d o m S e q u e n c e
N e g a tiv e B in o m ia l
N = 5
¿Cual es la probabilidad de encontrar un cluster con N CpGs y longitud X en
una distribución al azar? Binomial Negativa

Publicaciones sobre islas CpG
M. Gardiner-Garden and M. Frommer, CpG islands in vertebrate genomes. J. Mol. Biol., 196 (1987), pp. 261–282. D. Takai and P.A. Jones, Comprehensive analysis of CpG islands in human chromosomes 21 and 22. Proc. Natl. Acad. Sci. USA, 99 (2002), pp. 3740–3745. M. Hackenberg, C. Previti, P.L. Luque-Escamilla, P. Carpena, J. Martinez-Aroza and J.L. Oliver, CpGcluster: a distance-based algorithm for CpG-island detection. BMC Bioinformatics, 7 (2006), p. 446. Robert S. Illingworth, Adrian P. Bird CpG islands – ‘A rough guide’. FEBS letters (2009)

Detectar la metilación de DNA El problema: 1) Hibridación es insensible frente a la metilación no se puede usar chips de DNA 2) La PCR elimina la información acerca del estado de metilación Pre-tratamiento del DNA & lectura del estado de metilación

Bisulfito sódico & Secuenciación

Bisulfito sódico & Secuenciación
El protocolo: 1) Tratar el DNA con Bisulfito sódico una citosina no-metilada se desamina: citosina uracilo timina (PCR) una citosina metilada se mantiene
2) Secuenciar el DNA 3) Alinear las lecturas (los ‘reads’) al genoma un desemparejamiento (entre el read ) C/T indica no-metilación
un emparejamiento C/C indica metilación
Ventaja: • Se obtiene información de metilación para cada citosina y no solo valores medios para
una región como ocurre con muchos otros métodos • Se puede detectar la metilación en todos los contextos y no solo CpG
Reto: • Re-secuenciar un genoma entero • Alinear miles de millones de secuencias cortas (reads)

Tratamiento con Bisulfito
5' ...TUGTAAGCTGUGATGTCAGUGTAGTTCUGATG... 3'
3' ...AGUATTCGACGUTACAGTCGUATCAAGGUTAC... 5'
5' ...TCGTAAGCTGCGATGTCAGCGTAGTTCCGATG... 3' 3' ...AGCATTCGACGCTACAGTCGCATCAAGGCTAC... 5'
MethylC-Seq
5' ...TTGTAAGCTGTGATGTCAGTGTAGTTCTGATG... 3'
3' ...AGTATTCGACGTTACAGTCGTATCAAGGTTAC... 5'
BSW
BSC
BSW
BSC
Genomic Fragment
A
Denaturation
&
Bisulfite
modification
PCR I

Secuenciación
Métodos actuales Second Generation Sequencing
(Secuenciación masiva)
454 Pyrosequencing (PS)
Illumina Reversible Termination (RT)
SOLID Sequencing by Ligation (SBL)
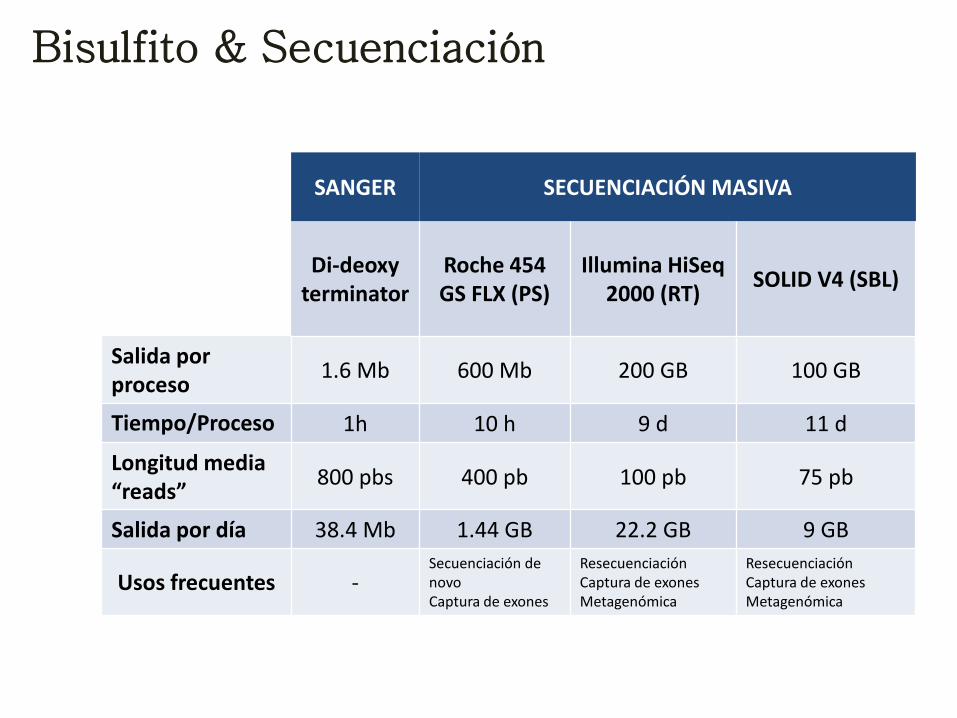
Bisulfito & Secuenciación
SANGER SECUENCIACIÓN MASIVA
Di-deoxy terminator
Roche 454 GS FLX (PS)
Illumina HiSeq 2000 (RT)
SOLID V4 (SBL)
Salida por proceso
1.6 Mb 600 Mb 200 GB 100 GB
Tiempo/Proceso 1h 10 h 9 d 11 d
Longitud media “reads”
800 pbs 400 pb 100 pb 75 pb
Salida por día 38.4 Mb 1.44 GB 22.2 GB 9 GB
Usos frecuentes - Secuenciación de novo Captura de exones
Resecuenciación Captura de exones Metagenómica
Resecuenciación Captura de exones Metagenómica

Muestra de DNA
Troceado
Selección
Reads secuenciados
Amplificació
n
Mapead
o
EXPERIMENTAL
COMPUTACIONAL
Genoma de referencia
Criterios de
calidad
Resecuenciación e interpretación
Diferencias: 1) Variación
2) Errores de secuenciación y
alineamiento
Alinear las lecturas

Obtener niveles de metilación

Context Start Watson Level Crick
level
Merged
Level
Watson
SNV fraction
Crick
SNV fraction Result
CG (+/-) 2 0.1 0.2 0.15 0 0.9 rejected
CTT (-) 4 - 0 - 0 - unMeth
CWG (+/-) 8 0.6 0.9 0.75 0 0 interMeth
CG (+/-) 11 0.2 0.1 0.15 0 0 unMeth
CAT (-) 13 - 0 0 1 - rejected
CWG (+/-) 17 0.3 0.1 0.2 0.2 0 unMeth
CG (+/-) 20 1 0.9 0.95 0 0 meth
CTA (-) 22 - 0.1 - 0 - unMeth
CCG (+) 27 0.2 - - - 0.2 unMeth
CG (+/-) 28 0.8 0.9 0.85 0.1 0 meth
CAT (-) 30 - 0 - 0.2 0 unMeth
Obtener niveles de metilación
Distinguir entre la acción del bisulfito y 1) Errores de secuenciación 2) SNV (Single Nucleotide
Variation) – un polimorfisomo con los alelos C/T seria detectado como una citosina no-metilado!!

Propiedades de la base de datos NGSmethDB
(http://bioinfo2.ugr.es/NGSmethDB/gbrowse)
• La base de datos NGSmethDB almacena datos de metilación a nivel de la citosina, generados por secuenciación masiva y conversión con bisulfito
• Actualmente contiene datos para 3 especies (humano, ratón y Arabidopsis) y un total de 22 tejidos únicos.
• Todos los datos se pueden bajar en formato texto. • No solo tiene datos del contexto CpG sino también de CWG (CAG/CTG) • El usuario puede elegir entre diferentes cuberturas (las veces que se ha secuenciado
una posición) • Para humano hay 1166274639, para ratón 69459481y para Arabidopsis 16321229
datos de metilación. • La base de datos cuenta con dos modos para obtener y analizar los datos. Primero,
un navegador (Genome Browser) para mostrar la metilación en el contexto de otras anotaciones como genes, islas CpG, contenido en G+C, etc. Segundo, varias pequeñas herramientas (php/Java) están disponibles para 1) buscar citosinas no metiladas, 2) detectar citosinas con metilación diferencial en un conjunto de tejidos, 3) obtener los niveles de metilación en una región dada, 4) analizar los niveles de metilación en la región promotora de los genes.
NGSmethDB

El navegador de NGSmethDB: La metilación se muestra mediante un gradiente de color: verde (no metilado) a rojo (metilado)
NGSmethDB
Stefanie Geisen, Guillermo Barturen, Ángel M. Alganza, Michael Hackenberg
and José L. Oliver. 2014. Nucleic Acids Research, Vol. 42, Database issue
D53–D59
http://bioinfo2.ugr.es/NGSmethDB/

Interfaz a las herramientas de NGSmethDB
NGSmethDB

















