Revisio ´n Vı ´a Wnt y esclerostina como nuevas dianas para la evaluacio ´n y el tratamiento de la osteoporosis Guillermo Alonso, Antonia Garcı ´a-Martı ´n y Manuel Mun ˜ oz-Torres * Unidad de Metabolismo O ´ seo, Servicio de Endocrinologı´a y Nutricio ´n, Hospital Universitario San Cecilio, Granada, Espan ˜a Descubrimiento y funciones de la vı ´a Wnt La vı ´a de sen ˜ alizacio ´n celular Wnt es esencial tanto para el desarrollo embrionario de diferentes especies animales estudiadas como para la regulacio ´n de diversos tejidos y o ´ rganos. Controla la proliferacio ´n y diferenciacio ´n celular y el mantenimiento de las ce ´ lulas madre, ası ´ como los movimientos celulares y el estable- cimiento de la polaridad tisular. Su disfuncio ´n se relaciona con la carcinoge ´ nesis y se ha implicado en la patogenia de diversas enfermedades degenerativas. Hoy en dı ´a constituye un objetivo potencial para la intervencio ´n terape ´ utica, sobre todo en los campos de la medicina regenerativa y de las ce ´ lulas madre 1 . Las bases para la investigacio ´n de esta vı ´a se iniciaron a finales de la de ´ cada de 1980 y principios de los 90 cuando se descubrio ´ que los genes de la mosca Drosophila wingless (Wg) y el rato ´n Int1 (Int1) eran reguladores de una gran familia de mole ´ culas de sen ˜ alizacio ´n conservada evolutivamente. El nombre Wnt deriva de la combi- nacio ´n de las palabras wingless e Int1 1 . Como puede deducirse de su participacio ´n en una amplia variedad de procesos biolo ´ gicos, la sen ˜ alizacio ´n de la vı ´a Wnt es inherentemente compleja. En te ´ rminos generales, esta compleji- dad se puede atribuir a 2 grandes aspectos. En primer lugar, tanto los ligandos como los receptores implicados pertenecen a grandes familias de proteı ´nas derivadas de mu ´ ltiples genes, lo que permite una amplia posibilidad en las interacciones ligando-receptor. En Med Clin (Barc). 2012;139(14):634–639 I N F O R M A C I O ´ N D E L A R T I ´ C U L O Historia del artı´culo: Recibido el 22 de enero de 2012 Aceptado el 1 de marzo de 2012 On-line el 19 de mayo de 2012 Palabras clave: Vı ´a Wnt Esclerostina Osteoporosis R E S U M E N El creciente conocimiento de la biologı ´a o ´ sea ha permitido la identificacio ´n de nuevas vı ´as intracelulares que participan en la regulacio ´n del remodelado y de la actividad de los osteoblastos. En este aspecto, la caracterizacio ´n de la vı ´a Wnt ha supuesto un gran avance por su implicacio ´n y relevancia en alteraciones del metabolismo mineral. El mejor conocimiento de estas vı ´as de sen ˜ alizacio ´n puede permitir desarrollar nuevos marcadores diagno ´ sticos, ası ´ como nuevos fa ´ rmacos para las enfermedades metabo ´ licas o ´ seas, donde a pesar del amplio arsenal terape ´ utico disponible au ´n persisten necesidades no cubiertas. En la presente revisio ´n realizamos una aproximacio ´n al descubrimiento y funciones de la vı ´a Wnt con especial atencio ´n a sus efectos o ´ seos. A continuacio ´ n, revisamos brevemente los datos principales acerca de su antagonista endo ´ geno, la esclerostina, por ser donde la investigacio ´n farmacolo ´ gica esta ´ ma ´s avanzada. ß 2012 Elsevier Espan ˜a, S.L. Todos los derechos reservados. Wnt pathway and sclerostin as new targets for assessment and treatment of osteoporosis Keywords: Wnt pathway Sclerostin Osteoporosis A B S T R A C T The increasing knowledge of bone biology has allowed the identification of new intracellular pathways involved in the regulation of remodelling and osteoblast activity. In this respect, the characterization of the Wnt pathway has been a breakthrough for its involvement and role in disorders of mineral metabolism. A better understanding of these signaling pathways may allow the development of new diagnostic markers and new drugs for metabolic bone disease, where despite extensive available therapies, unmet needs still persist. In this review, we make an approach to the discovery and functions of the Wnt pathway with a focus on bone effects. Next, we briefly review the main data about their endogenous antagonist, sclerostin, precisely where drug research is more advanced. ß 2012 Elsevier Espan ˜a, S.L. All rights reserved. * Autor para correspondencia. Correo electro ´nico: [email protected](M. Mun ˜ oz-Torres). www .elsevier .es/m ed icin ac linic a 0025-7753/$ – see front matter ß 2012 Elsevier Espan ˜a, S.L. Todos los derechos reservados. doi:10.1016/j.medcli.2012.03.006
Transcript
Med Clin (Barc). 2012;139(14):634–639
Revision
Vıa Wnt y esclerostina como nuevas dianas para la evaluacion y el tratamientode la osteoporosis
Guillermo Alonso, Antonia Garcıa-Martın y Manuel Munoz-Torres *
Unidad de Metabolismo Oseo, Servicio de Endocrinologıa y Nutricion, Hospital Universitario San Cecilio, Granada, Espana
I N F O R M A C I O N D E L A R T I C U L O
Historia del artıculo:
Recibido el 22 de enero de 2012
Aceptado el 1 de marzo de 2012
On-line el 19 de mayo de 2012
Palabras clave:
Vıa Wnt
Esclerostina
Osteoporosis
R E S U M E N
El creciente conocimiento de la biologıa osea ha permitido la identificacion de nuevas vıas intracelulares
que participan en la regulacion del remodelado y de la actividad de los osteoblastos. En este aspecto, la
caracterizacion de la vıa Wnt ha supuesto un gran avance por su implicacion y relevancia en alteraciones
del metabolismo mineral.
El mejor conocimiento de estas vıas de senalizacion puede permitir desarrollar nuevos marcadores
diagnosticos, ası como nuevos farmacos para las enfermedades metabolicas oseas, donde a pesar del
amplio arsenal terapeutico disponible aun persisten necesidades no cubiertas. En la presente revision
realizamos una aproximacion al descubrimiento y funciones de la vıa Wnt con especial atencion a sus
efectos oseos. A continuacion, revisamos brevemente los datos principales acerca de su antagonista
endogeno, la esclerostina, por ser donde la investigacion farmacologica esta mas avanzada.
� 2012 Elsevier Espana, S.L. Todos los derechos reservados.
Wnt pathway and sclerostin as new targets for assessment and treatment ofosteoporosis
Keywords:
Wnt pathway
Sclerostin
Osteoporosis
A B S T R A C T
The increasing knowledge of bone biology has allowed the identification of new intracellular pathways
involved in the regulation of remodelling and osteoblast activity. In this respect, the characterization of
the Wnt pathway has been a breakthrough for its involvement and role in disorders of mineral
metabolism.
A better understanding of these signaling pathways may allow the development of new diagnostic
markers and new drugs for metabolic bone disease, where despite extensive available therapies, unmet
needs still persist. In this review, we make an approach to the discovery and functions of the Wnt
pathway with a focus on bone effects. Next, we briefly review the main data about their endogenous
antagonist, sclerostin, precisely where drug research is more advanced.
� 2012 Elsevier Espana, S.L. All rights reserved.
www .e lsev ier .es /m ed i c in ac l in ic a
Descubrimiento y funciones de la vıa Wnt
La vıa de senalizacion celular Wnt es esencial tanto para eldesarrollo embrionario de diferentes especies animales estudiadascomo para la regulacion de diversos tejidos y organos. Controla laproliferacion y diferenciacion celular y el mantenimiento de lascelulas madre, ası como los movimientos celulares y el estable-cimiento de la polaridad tisular. Su disfuncion se relaciona con lacarcinogenesis y se ha implicado en la patogenia de diversasenfermedades degenerativas. Hoy en dıa constituye un objetivo
0025-7753/$ – see front matter � 2012 Elsevier Espana, S.L. Todos los derechos reserv
doi:10.1016/j.medcli.2012.03.006
potencial para la intervencion terapeutica, sobre todo en loscampos de la medicina regenerativa y de las celulas madre1.
Las bases para la investigacion de esta vıa se iniciaron a finalesde la decada de 1980 y principios de los 90 cuando se descubrio quelos genes de la mosca Drosophila wingless (Wg) y el raton Int1 (Int1)eran reguladores de una gran familia de moleculas de senalizacionconservada evolutivamente. El nombre Wnt deriva de la combi-nacion de las palabras wingless e Int11.
Como puede deducirse de su participacion en una ampliavariedad de procesos biologicos, la senalizacion de la vıa Wnt esinherentemente compleja. En terminos generales, esta compleji-dad se puede atribuir a 2 grandes aspectos. En primer lugar, tantolos ligandos como los receptores implicados pertenecen a grandesfamilias de proteınas derivadas de multiples genes, lo que permiteuna amplia posibilidad en las interacciones ligando-receptor. En
G. Alonso et al / Med Clin (Barc). 2012;139(14):634–639 635
segundo lugar, estas interacciones producen diversas respuestasintracelulares interrelacionadas entre sı2.
Efectos oseos
Los datos disponibles indican que las proteınas Wnt ejercendiversas funciones en la genesis del esqueleto, que van desde ladefinicion de patrones del esqueleto embrionario hasta elremodelado del esqueleto adulto, pasando por el desarrollo delesqueleto fetal. En el esqueleto adulto, las celulas madremesenquimales se diferencian a celulas osteoprogenitoras bajocondiciones adecuadas. La vıa canonica de senalizacion Wntfavorece la lınea celular osteoblastogenica mediante el estımulo dela proliferacion, diferenciacion y maduracion del osteoblasto atraves de la induccion de los factores de transcripcion osteoge-nicos. Ademas, inhibe la diferenciacion a condrocito y del adipocitodesde las celulas progenitoras. Asimismo, regula la apoptosis tantode los osteoprogenitores indiferenciados como de los osteoblastosdiferenciados3 (fig. 1).
Los datos recientes in vitro e in vivo confirman que las proteınasWnt inducen la produccion de osteoprotegerina (OPG) por elosteoblasto, inhibiendo por tanto la diferenciacion del osteoclastoy la resorcion osea. Por tanto, la vıa Wnt parece controlarindirectamente la osteoclastogenesis a traves de sus efectos sobrelos osteoblastos4,5. Las observaciones sobre la relacion con elsistema OPG/RANKL no solo pueden ser relevantes en laosteoporosis, sino tambien en las lesiones oseas asociadas atumores y en diversas enfermedades osteoarticulares.
Vıa intracelular canonica
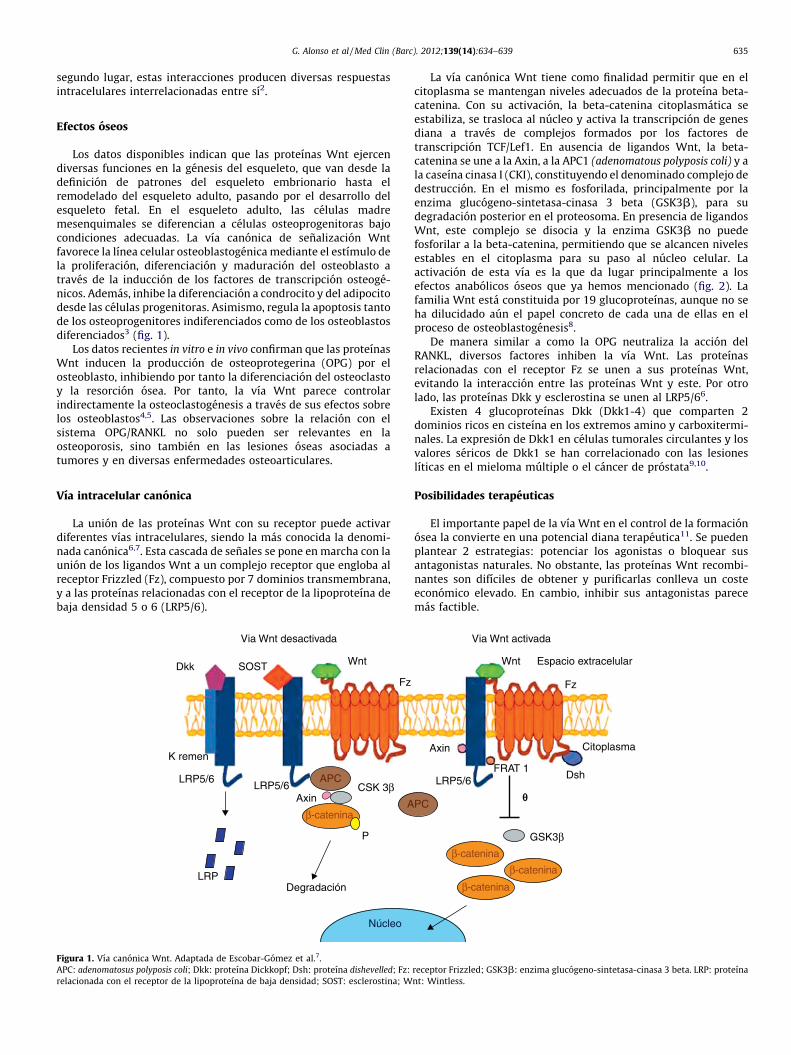
La union de las proteınas Wnt con su receptor puede activardiferentes vıas intracelulares, siendo la mas conocida la denomi-nada canonica6,7. Esta cascada de senales se pone en marcha con launion de los ligandos Wnt a un complejo receptor que engloba alreceptor Frizzled (Fz), compuesto por 7 dominios transmembrana,y a las proteınas relacionadas con el receptor de la lipoproteına debaja densidad 5 o 6 (LRP5/6).
Dkk SOST
K remen
LRP5/6LRP5/6
Fz
Wnt
Via Wnt desactivada
Axin
Degradación
Núcleo
β-catenina
P
CSK 3βA
APC
LRP
Figura 1. Vıa canonica Wnt. Adaptada de Escobar-Gomez et al.7.
relacionada con el receptor de la lipoproteına de baja densidad; SOST: esclerostina; W
La vıa canonica Wnt tiene como finalidad permitir que en elcitoplasma se mantengan niveles adecuados de la proteına beta-catenina. Con su activacion, la beta-catenina citoplasmatica seestabiliza, se trasloca al nucleo y activa la transcripcion de genesdiana a traves de complejos formados por los factores detranscripcion TCF/Lef1. En ausencia de ligandos Wnt, la beta-catenina se une a la Axin, a la APC1 (adenomatous polyposis coli) y ala caseına cinasa I (CKI), constituyendo el denominado complejo dedestruccion. En el mismo es fosforilada, principalmente por laenzima glucogeno-sintetasa-cinasa 3 beta (GSK3b), para sudegradacion posterior en el proteosoma. En presencia de ligandosWnt, este complejo se disocia y la enzima GSK3b no puedefosforilar a la beta-catenina, permitiendo que se alcancen nivelesestables en el citoplasma para su paso al nucleo celular. Laactivacion de esta vıa es la que da lugar principalmente a losefectos anabolicos oseos que ya hemos mencionado (fig. 2). Lafamilia Wnt esta constituida por 19 glucoproteınas, aunque no seha dilucidado aun el papel concreto de cada una de ellas en elproceso de osteoblastogenesis8.
De manera similar a como la OPG neutraliza la accion delRANKL, diversos factores inhiben la vıa Wnt. Las proteınasrelacionadas con el receptor Fz se unen a sus proteınas Wnt,evitando la interaccion entre las proteınas Wnt y este. Por otrolado, las proteınas Dkk y esclerostina se unen al LRP5/66.
Existen 4 glucoproteınas Dkk (Dkk1-4) que comparten 2dominios ricos en cisteına en los extremos amino y carboxitermi-nales. La expresion de Dkk1 en celulas tumorales circulantes y losvalores sericos de Dkk1 se han correlacionado con las lesioneslıticas en el mieloma multiple o el cancer de prostata9,10.
Posibilidades terapeuticas
El importante papel de la vıa Wnt en el control de la formacionosea la convierte en una potencial diana terapeutica11. Se puedenplantear 2 estrategias: potenciar los agonistas o bloquear susantagonistas naturales. No obstante, las proteınas Wnt recombi-nantes son difıciles de obtener y purificarlas conlleva un costeeconomico elevado. En cambio, inhibir sus antagonistas parecemas factible.
G. Alonso et al / Med Clin (Barc). 2012;139(14):634–639636
En la actualidad se estan investigando pequenas moleculas queinactivan enzimas intracelulares (principalmente GSK3b), con loque se reduce la actividad de la beta-catenina. El litio es uninhibidor conocido de la enzima GSK3b que estimula ladiferenciacion osteoblastica in vitro y la regeneracion osea in vivo.Ası, en los pacientes con trastorno bipolar tratados con litio losestudios epidemiologicos sugieren menor riesgo de fractura12.Aunque existen moleculas especıficas inhibidoras de la GSK3b quehan demostrado aumentar la masa osea, aun deben investigarse lospotenciales efectos de carcinogenesis debido a la ubicuidad de lavıa Wnt y de esta enzima13.
Los anticuerpos neutralizantes frente a Dkk1 han mostradoefectos positivos sobre la densidad mineral osea en modelosanimales, aunque los estudios aun se encuentran en fasestempranas y la expresion de Dkk1 en multiples tejidos podrıarestarle eficacia y seguridad a nivel oseo14. Los estudios sobreanticuerpos frente a esclerostina son los que se encuentran en fasesmas avanzadas con prometedores resultados15–17, como se exponemas adelante.
Esclerostina
Descubrimiento y sıntesis
A partir del estudio genetico y molecular de 2 displasiasesclerosantes poco frecuentes, la esclerosteosis y la enfermedad deVan Buchem, se describio en 2001 la proteına esclerostina.
La esclerosteosis, una enfermedad autosomica recesiva descritapor primera vez en 1958, se caracteriza por un progresivoengrosamiento oseo con sindactilia, talla alta y agrandamientodel craneo. La enfermedad de Van Buchem es otro defectoautosomico recesivo similar, de gravedad menor y en la que nose presentan los hallazgos de sindactilia ni talla alta18. En ambosprocesos se observa una importante hiperostosis endostal conincremento de osteoblastos activos y osteoide adecuadamentemineralizado. A pesar de las marcadas anomalıas oseas, es notableel hecho de que los individuos afectados no presentan un riesgoaumentado de fracturas19.
El defecto genetico que produce la esclerosteosis ha sidoidentificado como mutaciones con perdida de funcion del gendenominado SOST, localizado en el cromosoma 17q12-21 yencargado de la codificacion de la proteına esclerostina20. En laenfermedad de Van Buchem se produce una delecion de este gen. Elresultado es la ausencia o la disminucion de la expresion genica deesclerostina, respectivamente, lo que condiciona diversos fenoti-pos oseos en funcion de la densidad mineral osea21. Por otro lado,las mutaciones en el gen LRP5 conllevan tambien aumento de lamasa osea.
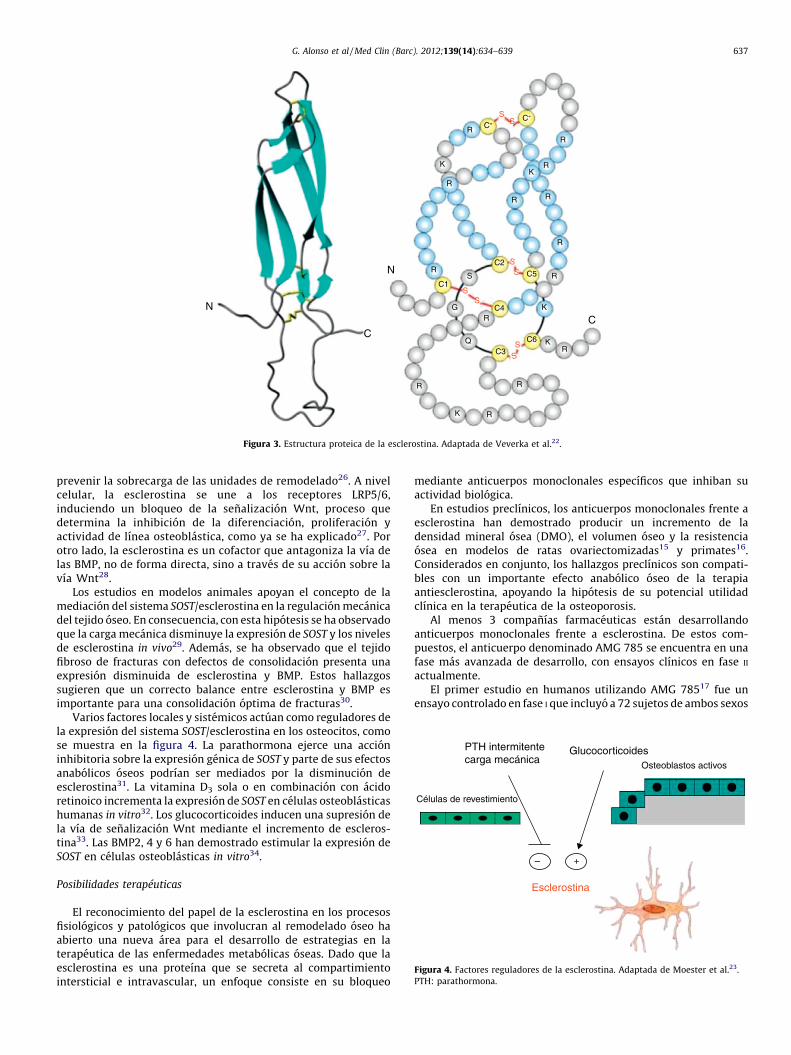
La esclerostina es una glucoproteına de 190 aminoacidosmiembro de la familia proteica DAN, a la que tambien pertenecenlas proteınas que se unen e inhiben a las BMP. Asimismo, compartesimilitudes estructurales con otro antagonista de la vıa Wnt,Wise22 (fig. 3).
En ratas adultas y en tejido oseo humano, la expresion deesclerostina esta restringida a los osteocitos, mientras que no seexpresa en osteoblastos, osteoclastos ni en las celulas derevestimiento. Tambien se ha detectado su expresion genica enrinon, hıgado, placenta y cartılago, aunque no se ha aislado laproteına como tal23. El significado fisiologico de este hallazgo estaaun por determinar.
La esclerostina se libera a la circulacion, siendo posible ladeterminacion de sus valores sericos gracias al desarrollo de variosensayos24. Al mismo tiempo, existe una fuerte correlacion entre elcontenido oseo de esclerostina y los valores circulantes, lo queindica la produccion exclusiva de esta proteına en el hueso y lafiabilidad de su medicion en suero25.
De esta forma, en la actualidad cada vez se publican mastrabajos clınicos centrados en el analisis de la concentracion sericade esclerostina en diversas situaciones clınicas, los factores queinfluyen en la misma, su asociacion con el metabolismo oseo o loscambios inducidos por los tratamientos para la osteoporosis.
Funciones y regulacion
La expresion de esclerostina por los osteocitos al inicio delproceso de mineralizacion de la sustancia osteoide actuarıa comosenal de retroalimentacion negativa sobre los osteoblastos para
GlucocorticoidesPTH intermitentecarga mecánica
Esclerostina
Osteoblastos activos
Células de revestimiento
– +
Figura 4. Factores reguladores de la esclerostina. Adaptada de Moester et al.23.
PTH: parathormona.
N
KK
R C-C-
R
R
R R
R
R
R
R
K
C3
C6
R
C2
C4
C5
S
SS
S
SS
S
S
SC1
R R
R
RK
K
Q
G
CC
N
Figura 3. Estructura proteica de la esclerostina. Adaptada de Veverka et al.22.
G. Alonso et al / Med Clin (Barc). 2012;139(14):634–639 637
prevenir la sobrecarga de las unidades de remodelado26. A nivelcelular, la esclerostina se une a los receptores LRP5/6,induciendo un bloqueo de la senalizacion Wnt, proceso quedetermina la inhibicion de la diferenciacion, proliferacion yactividad de lınea osteoblastica, como ya se ha explicado27. Porotro lado, la esclerostina es un cofactor que antagoniza la vıa delas BMP, no de forma directa, sino a traves de su accion sobre lavıa Wnt28.
Los estudios en modelos animales apoyan el concepto de lamediacion del sistema SOST/esclerostina en la regulacion mecanicadel tejido oseo. En consecuencia, con esta hipotesis se ha observadoque la carga mecanica disminuye la expresion de SOST y los nivelesde esclerostina in vivo29. Ademas, se ha observado que el tejidofibroso de fracturas con defectos de consolidacion presenta unaexpresion disminuida de esclerostina y BMP. Estos hallazgossugieren que un correcto balance entre esclerostina y BMP esimportante para una consolidacion optima de fracturas30.
Varios factores locales y sistemicos actuan como reguladores dela expresion del sistema SOST/esclerostina en los osteocitos, comose muestra en la figura 4. La parathormona ejerce una accioninhibitoria sobre la expresion genica de SOST y parte de sus efectosanabolicos oseos podrıan ser mediados por la disminucion deesclerostina31. La vitamina D3 sola o en combinacion con acidoretinoico incrementa la expresion de SOST en celulas osteoblasticashumanas in vitro32. Los glucocorticoides inducen una supresion dela vıa de senalizacion Wnt mediante el incremento de escleros-tina33. Las BMP2, 4 y 6 han demostrado estimular la expresion deSOST en celulas osteoblasticas in vitro34.
Posibilidades terapeuticas
El reconocimiento del papel de la esclerostina en los procesosfisiologicos y patologicos que involucran al remodelado oseo haabierto una nueva area para el desarrollo de estrategias en laterapeutica de las enfermedades metabolicas oseas. Dado que laesclerostina es una proteına que se secreta al compartimientointersticial e intravascular, un enfoque consiste en su bloqueo
mediante anticuerpos monoclonales especıficos que inhiban suactividad biologica.
En estudios preclınicos, los anticuerpos monoclonales frente aesclerostina han demostrado producir un incremento de ladensidad mineral osea (DMO), el volumen oseo y la resistenciaosea en modelos de ratas ovariectomizadas15 y primates16.Considerados en conjunto, los hallazgos preclınicos son compati-bles con un importante efecto anabolico oseo de la terapiaantiesclerostina, apoyando la hipotesis de su potencial utilidadclınica en la terapeutica de la osteoporosis.
Al menos 3 companıas farmaceuticas estan desarrollandoanticuerpos monoclonales frente a esclerostina. De estos com-puestos, el anticuerpo denominado AMG 785 se encuentra en unafase mas avanzada de desarrollo, con ensayos clınicos en fase II
actualmente.El primer estudio en humanos utilizando AMG 78517 fue un
ensayo controlado en fase I que incluyo a 72 sujetos de ambos sexos
G. Alonso et al / Med Clin (Barc). 2012;139(14):634–639638
aleatorizados a recibir el producto de investigacion (n = 54) oplacebo (n = 18). Se administraron diferentes dosis por vıasubcutanea (SC) a 56 individuos y por vıa intravenosa (IV) a 16participantes. Tras la administracion de una dosis simple de AMG785, hubo un incremento dependiente de la dosis de marcadoresde formacion osea (P1NP, BSAP y osteocalcina) y una disminucionde marcadores de resorcion (CTx serico). El analisis del patron dedichos cambios sugirio un desacoplamiento entre la actividadosteoblastica y la osteoclastica. El mayor incremento de DMO fueobservado en el dıa 85 tanto para la vıa SC (incremento en la DMOlumbar del 5,3% y femoral total del 2,8%; p < 0,01) como IV (lumbar5,2%, p < 0,01; femoral total 1,1%, p > 0,05). La mayorıa de losefectos adversos fueron considerados leves y ninguno resulto endiscontinuacion, siendo los mas frecuentemente descritos: eritemaen el sitio de inyeccion, dolor lumbar, cefalea, estrenimiento,artralgias y mareos. Ademas, se observo una disminucion leve,transitoria y asintomatica del calcio ionico serico (alrededor del 4%por debajo del valor basal). Tomando como referencia detratamiento osteoanabolico a teriparatida, los hallazgos prelimi-nares con AMG 785 mostraron un incremento de la DMO a 3 mesessimilar o superior a los observados tras 6 meses con teriparatida. Elpatron de cambios en marcadores de remodelado oseo tambienparece ser diferente. Ası, AMG 785 producirıa un desacoplamientoformacion-resorcion, con un incremento en la formacion queocurre al mismo tiempo que disminuye la resorcion osea. Estosugiere una importante ventana anabolica que posibilitarıa unamayor respuesta en terminos de ganancia de DMO y mayorduracion de la terapia. No obstante, son necesarios mas estudiospara evaluar los aspectos de mayor trascendencia clınica, como laseguridad a largo plazo, el impacto sobre la incidencia de fracturas,la consolidacion de estas y la reparacion osea tras intervencionesquirurgicas.
De esta manera, los datos disponibles en la actualidad abrennuevas y prometedoras perspectivas en la evaluacion y terapeuticade la osteoporosis35 mediante el nuevo paradigma del bloqueo dela esclerostina mediante anticuerpos monoclonales especıficos.
No obstante, son necesarios mas estudios para evaluar losaspectos de mayor trascendencia clınica, como la seguridad a largoplazo, el impacto sobre la incidencia y consolidacion de lasfracturas y la reparacion osea tras intervenciones quirurgicas.
Conclusiones
La vıa de senalizacion Wnt constituye un mecanismo esencialde la regulacion del remodelado oseo, implica el correctofuncionamiento de diversos factores conectados entre sı, y escapaz de ejercer un control global sobre el osteoblasto, favore-ciendo su proliferacion, diferenciacion o apoptosis.
Los tratamientos actuales para la osteoporosis estan casiexclusivamente basados en un efecto antirresortivo36, por lo queexiste una necesidad real para terapias alternativas basadas en laestimulacion de vıas anabolicas en el hueso.
El hallazgo de proteınas antagonistas de la vıa Wnt, como laesclerostina, esta permitiendo descubrir nuevas dianas terapeu-ticas que ejerzan un efecto anabolico en el tejido oseo, al mismotiempo que no alteren su funcion biomecanica fisiologica.
Conflicto de intereses
Los autores declaran no tener ningun conflicto de intereses.
Bibliografıa
1. Van Amerongen R, Nusse R. Towards an integrated view of Wnt signaling indevelopment. Development. 2009;136:3205–14.
2. Gordon MD, Nusse R. Wnt signaling: multiple pathways, multiple receptors andmultiple transcription factors. J Biol Chem. 2006;281:22429–33.
3. Baron R, Rawadi G. Wnt signaling and the regulation of bone mass. CurrOsteoporos Rep. 2007;5:73–80.
4. Glass DA, Bialek P, Ahn JD, Starbuck M, Patel MS, Clevers H, et al. Canonical Wntsignaling in differentiated osteoblasts controls osteoclast differentiation. DevCell. 2005;8:751–64.
5. Kamiya N, Ye L, Kobayashi T, Mochida Y, Yamauchi M, Kronenberg HM, et al.BMP signaling negatively regulates bone mass through sclerostin by the cano-nical Wnt pathway. Development. 2008;135:3801–11.
6. Baron R, Rawadi G. Targeting the Wnt/beta-catenin pathway to regulate boneformation in the adult skeleton. Endocrinology. 2007;148:2635–43.
7. Escobar-Gomez F, Jodar E, Hawkins F. Receptor Wnt: fisiologıa, fisiopatologıa ypotenciales nuevas dianas terapeuticas. Rev Esp Enferm Metab Oseas.2009;18:39–44.
8. Krishnan V, Bryant HU, McDougald OA. Regulation of bone mass by Wntsignalling. J Clin Invest. 2006;116:1202–9.
9. Tian E, Zhan F, Walker R, Rasmussen E, Ma Y, Barlogie B, et al. The role of theWnt-signaling antagonist DKK1 in the development of osteolytic lesions inmultiple myeloma. N Engl J Med. 2003;349:2483–94.
10. Hall CL, Bafico A, Dai J, Aaronson SA, Keller ET. Prostate cancer cells promoteosteoblastic bone metastases through Wnts. Cancer Res. 2005;65:7554–60.
11. Hoeppner LH, Secreto FJ, Westendorf JJ. Wnt signaling as a therapeutic target forbone diseases. Expert Opin Ther Targets. 2009;13:485–96.
12. Vestergaard P, Rejnmark L, Mosekilde L. Reduced relative risk of fracturesamong users of lithium. Calcif Tissue Int. 2005;77:1–8.
13. Kulkarni NH, Onyia JE, Zeng Q, Tian X, Liu M, Halladay DL, et al. Orallybioavailable GSK-3alpha/beta dual inhibitor increases markers of cellulardifferentiation in vitro and bone mass in vivo. J Bone Miner Res. 2006;21:910–20.
14. Glantschnig H, Scott K, Hampton R, Wei N, McCracken P, Nantermet P, et al. Arate-limiting role for Dickkopf-1 in bone formation and the remediation of boneloss in mouse and primate models of postmenopausal osteoporosis by anexperimental therapeutic antibody. J Pharmacol Exp Ther. 2011;338:568–78.
15. Li X, Ominsky MS, Warmington KS, Morony S, Gong J, Cao J, et al. Sclerostinantibody treatment increases bone formation, bone mass, and bone strength ina rat model of postmenopausal osteoporosis. J Bone Miner Res. 2009;24:578–88.
16. Ominsky MS, Vlasseros F, Jolette J, Smith SY, Stouch B, Doellgast G, et al. Twodoses of sclerostin antibody in cynomolgus monkeys increases bone formation,bone mineral density and bone strength. J Bone Miner Res. 2010;25:948–59.
17. Padhi D, Jang G, Stouch B, Fang L, Posvar E. Single-dose, placebo-controlled,randomized study of AMG 785, a sclerostin monoclonal antibody. J Bone MinerRes. 2011;26:19–26.
18. Staehling-Hampton K, Proll S, Paeper BW, Zhao L, Charmley P, Brown A, et al. A52-kb deletion in the SOST-MEOX1 intergenic region on 17q12-q21 is asso-ciated with van Buchem disease in the Dutch population. Am J Med Genet.2002;110:144–52.
19. Hamersma H, Gardner J, Beighton P. The natural history of sclerosteosis. ClinGenet. 2003;63:192–7.
20. Brunkow ME, Gardner JC, van Ness J, Paeper BW, Kovacevich BR, Proll S, et al.Bone dysplasia sclerosteosis results from loss of the SOST gene product, a novelcystine knot-containing protein. Am J Hum Genet. 2001;68:577–89.
21. Balemans W, Cleiren E, Siebers U, Horst J, van Hul W. A generalized skeletalhyperostosis in two siblings caused by a novel mutation in the SOST gene. Bone.2005;36:943–7.
22. Veverka V, Henry AJ, Slocombe PM, Ventom A, Mulloy B, Muskett FW, et al.Characterization of the structural features and interactions of sclerostin: mole-cular insight into a key regulator of Wnt-mediated bone formation. J Biol Chem.2009;284:10890–900.
23. Moester MJ, Papapoulos SE, Lowik CW, van Bezooijen RL. Sclerostin: currentknowledge and future perspectives. Calcif Tissue Int. 2010;87:99–107.
24. McNulty M, Singh RJ, Li X, Bergstralh EJ, Kumar R. Determination of serum andplasma sclerostin concentrations by enzyme-linked immunoassays. J ClinEndocrinol Metab. 2011;96:1156–62.
25. Drake MT, Srinivasan B, Modder UI, Peterson JM, McCready LK, Riggs BL, et al.Effects of parathyroid hormone treatment on circulating sclerostin levels inpostmenopausal women. J Clin Endocrinol Metab. 2010;95:5056–62.
26. Poole KE, van Bezooijen RL, Loveridge N, Hamersma H, Papapoulos SE, LowikCW, et al. Sclerostin is a delayed secreted product of osteocytes that inhibitsbone formation. FASEB J. 2005;19:1842–4.
27. Li X, Zhang Y, Kang H, Liu W, Liu P, Zhang J, et al. Sclerostin binds to LRP5/6 andantagonizes canonical Wnt signaling. J Biol Chem. 2005;280:19883–7.
28. Van Bezooijen RL, Svensson JP, Eefting D, Visser A, van der Horst G, Karperien M,et al. Wnt but not BMP signaling is involved in the inhibitory action of sclerostinon BMP-stimulated bone formation. J Bone Miner Res. 2007;22:19–28.
29. Robling AG, Niziolek PJ, Baldridge LA, Condon KW, Allen MR, Alam I, et al.Mechanical stimulation of bone in vivo reduces osteocyte expression of Sost/sclerostin. J Biol Chem. 2008;283:5866–75.
30. Niikura T, Hak DJ, Reddi AH. Global gene profiling reveals a downregulation ofBMP gene expression in experimental atrophic nonunions compared to stan-dard healing fractures. J Orthop Res. 2006;24:1463–71.
31. O’Brien CA, Plotkin LI, Galli C, Goellner JJ, Gortazar AR, Allen MR, et al. Control ofbone mass and remodeling by PTH receptor signaling in osteocytes. PLoS One.2008;3:e2942.
32. Sutherland MK, Geoghegan JC, Yu C, Winkler DG, Latham JA. Unique regulationof SOST, the sclerosteosis gene, by BMPs and steroid hormones in humanosteoblasts. Bone. 2005;35:448–54.
G. Alonso et al / Med Clin (Barc). 2012;139(14):634–639 639
33. Yao W, Cheng Z, Pham A, Busse C, Zimmermann EA, Ritchie RO, et al. Gluco-corticoid-induced bone loss in mice can be reversed by the actions of para-thyroid hormone and risedronate on different pathways for bone formation andmineralization. Arthritis Rheum. 2008;58:3485–97.
34. Ohyama Y, Nifuji A, Maeda Y, Amagasa T, Noda M. Spaciotemporal associationand bone morphogenetic protein regulation of sclerostin and osterix expressionduring embryonic osteogenesis. Endocrinology. 2004;145:4685–92.
35. Vazquez M. Osteoporosis: la crisis de un paradigma. Med Clin (Barc).2010;134:206–7.
36. Kanterewicz E, Puigoriol E, Peris P, del Rıo L, Rosique P, Yanez A, et al.Relacion del telopeptido carboxiterminal del colageno I serico con ladensidad mineral osea y el consumo de farmacos en mujeres posmenopau-sicas. Datos preliminares del estudio FRODOS. Med Clin (Barc). 2009;133:609–14.