

APORTACIONES AL ESTUDIO
ULTRAESTRUCTURAL DEL SISTEMA VISUAL
Natalia Pérez Losantos
Directora: Mª José Luesma Bartolomé
Grado de Óptica y Optometría 2014 - 2015


ÍNDICE
I. Justificación
II. Introducción
III. Materiales y métodos
a) Microscopio óptico
1. Disección y fijación
2. Inclusión
3. Corte
4. Tinción
b) Microscopio electrónico
1. Fijación
2. Inclusión
3. Corte
4. Contraste o tinción
IV. Introducción general de la estructura del globo ocular
a) Túnica externa o fibrosa
1. Esclerótica
2. Córnea
b) Túnica media o vascular. Úvea
c) Túnica interna o nerviosa. Retina
V. Resultados. Aportaciones al estudio ultraestructural del sistema visual
a) Coroides
b) Cuerpo ciliar
1. Epitelio ciliar
2. Estroma vascular
3. Músculo ciliar
VI. Conclusiones
VII. Bibliografía
ANEXOS

I. Justificación
La ley 44/2003 de Ordenación de las Profesiones Sanitarias, reconoce al Graduado/a en Óptica
y Optometría como un/a profesional sanitario/a, la misma Ordenación establece como
funciones del Óptico – optometrista las actividades dirigidas a la detección de los defectos de
la refracción ocular, a través de su medida instrumental, a la utilización de medidas de
reeducación, prevención e higiene visual, y a la adaptación, verificación y control de las ayudas
ópticas, todo esto hace de la materia Anatomía e Histología Ocular una disciplina
imprescindible para la identificación de las características anatomohistológicas de normalidad
y salud del sistema visual.
La necesidad del presente trabajo surge porque, tanto la Anatomía como la Histología, son dos
disciplinas altamente descriptivas y si bien la bibliografía en ambos campos es extensa, se
reduce mucho cuando lo acotamos al campo del sistema visual, especialización imprescindible
para los Graduados en Óptica y Optometría. Es por ello que consideramos necesaria una
recopilación de imágenes con fines didácticos específicos en la materia. Nuestro estudio se
centrará en concreto en el cuerpo ciliar, parte integrante de la túnica vascular o úvea.
La microscopía óptica y la electrónica son dos herramientas de gran valor que nos permiten el
estudio de las estructuras microscópicas y ultraestructurales del globo ocular y de los anejos.
II. Introducción
Los sistemas de estudio morfológicos de células y tejidos empleados son la microscopía óptica
y la microscopía electrónica.
El microscopio es un instrumento óptico que, como su nombre indica (del griego mikrós,
“pequeño”, y skopein, “examinar”), permite ver objetos que por su pequeño tamaño escapan a
la percepción del ojo humano.
Los microscopios ópticos o fotónicos se dividen en los siguientes tipos: microscopio simple,
estereoscópico o lupa y microscopio compuesto, también llamado común.
El microscopio simple está constituido por una lente o sistema de lentes convergentes
dispuestas de tal modo que suministran una imagen virtual, derecha y mayor que el objeto;
éste se encuentra situado entre la lente y su foco.
Las lupas amplían 2, 4, 8, 10 ó 20 veces, o algo más, la imagen del objeto observado. Existen
lupas monoculares y binoculares, montadas en un soporte cuya lente se acerca o aleja del
objeto, depositado sobre una platina, mediante un mecanismo de cremallera accionado por un
piñón. Sus aplicaciones son bastante limitadas; suelen utilizarse para la disección de pequeños
animales o para la disociación de piezas histológicas.
En el microscopio óptico compuesto se combinan los poderes amplificantes de dos lentes o
sistemas de lentes, ambos convergentes, que se encuentran colocados en los extremos de un
tubo: el objetivo, situado cerca del objeto que se observa, y el ocular, colocado cerca del ojo.
1

Con este instrumento se logran aumentos de 25, 50, 100, 1.000 y hasta 1.500 veces según la
combinación de lentes y la longitud de onda que se emplea para su iluminación. La imagen
resultante es virtual e invertida.
Dicho microscopio posee una parte mecánica y otra óptica.
La parte mecánica está constituida por el estátivo, o montura del microscopio, y consta de pie,
columna, platina, tubo y revólver.
La parte óptica comprende los objetivos, los oculares y el aparato de iluminación.
El objetivo está compuesto por un sistema de lentes convergentes montadas en un tubo
metálico que constituye el soporte del objetivo. Este sistema proporciona una imagen real,
aumentada e invertida, del objeto que se observa.
Además del aumento, una propiedad importante de un microscopio es su poder de resolución
(o de separación), es decir, la capacidad de mostrar distintos y separados dos puntos muy
próximos. Por consiguiente, cuanto mayor sea el poder de resolución, mayor será la definición
de un objeto.
El ocular consta de dos lentes o sistemas de ellas que se encuentran montadas en los extremos
de un tubo adaptado a la parte superior del microscopio. El ocular recoge la imagen
suministrada por el objetivo transformándola en una imagen virtual, aumentada y derecha. La
lente superior se denomina lente ocular, y la inferior, colectora o lente de campo.
El sistema de iluminación de un microscopio es también de considerable importancia, sobre
todo cuando se utilizan grandes aumentos.
A diferencia del microscopio óptico, el microscopio electrónico utiliza como fuente de luz un
haz de electrones de longitud de onda 100.000 veces inferior a la de la luz empleada
habitualmente. Este instrumento permite obtener aumentos muy superiores a los del
microscopio óptico y con un límite de resolución de hasta 5 A. además, permite estudiar
estructuras complejas o niveles de organización de la materia viva y de cada uno de sus
componentes que no pueden ser estudiados con el microscopio óptico.
Las partes del microscopio electrónico son: tubo, pantalla de proyección, filamento (situado en
la extremidad superior del tubo), lentes, portamuestras, brazo articulado para manejo de
muestras y cámara fotográfica o de TV.
Para llevar a cabo el funcionamiento del microscopio electrónico hay que conseguir un haz de
electrones a partir de un filamento metálico de tungsteno.
El análisis definitivo de la preparación se hace a partir de las fotografías realizadas con
aparición de una imagen en blanco y negro1, 2.
El principio de la microscopía electrónica es muy similar al de la microscopia de luz y se han
desarrollado dos principales técnicas:
2

1. Microscopía electrónica de transmisión.
Se observa a través del espécimen (trans – iluminación). El espécimen se corta en
láminas ultrafinas (en el orden de nanómetros) que se colocan en una rejilla de cobre, la
cual es bombardeada con un haz de electrones enfocado.
Lo que podemos ver es la estructura interna.
2. Microscopía electrónica de barrido.
Se observa la superficie de un espécimen sólido (epi – iluminación). Se producen
imágenes muy interesantes en 3D gracias a una mayor profundidad de campo.
Lo que podemos ver es la superficie.
Nosotros emplearemos el microscopio de transmisión.
A continuación, mostraremos un cuadro comparativo entre el microscopio óptico y el
microscopio electrónico3, 4.
MICROSCOPIO ÓPTICO
MICROSCOPIO ELECTRÓNICO
Características: 3. Permite aumentos desde 25 a 1.500
veces 4. El poder de resolución es de 0.2 μm 5. La muestra es atravesada por la luz 6. Las lentes son de vidrio 7. Los cortes son muy finos y se
obtienen con un micrótomo
Características:
Permite aumentos superiores a 10.000, 20.000 y excepcionalmente 500.000
El poder de resolución es de unos 3 a 10 A
La muestra es atravesada por un haz de electrones emitidos por un filamento de tungsteno
Las “lentes” son campos magnéticos
Los cortes son ultra finos y se obtienen con un ultramicrotono
La imagen se genera sobre una pantalla fluorescente
Ventajas:
Se pueden observar células vivas
Se pueden ver células enteras
Ventajas:
Se puede ver la Ultraestructura de la célula en los cortes de los orgánulos
Inconvenientes:
Los análisis no pueden ser muy detallados y profundos
Inconvenientes:
No permite observar células vivas
No se pueden realizar visiones de conjunto
Unidades de medida:
El micrómetro (μm)
Unidades de medida:
La unidad oficial es el nanómetro (nm) pero es más empleado el angstrom (A)
3

III. Materiales y métodos
A pesar de la disparidad de tamaño, las similitudes en la estructura anatómica de los ojos
humanos y de ratón son mucho mayores que las diferencias. El desarrollo y la estructura
adulta de la córnea, segmento anterior, y el aparato de salida del humor acuoso son muy
similares en el ratón y en el humano, excepto por el menor tamaño relativo del músculo ciliar y
el tamaño relativo más grande de la lente5-7. Además, aunque la retina del ratón carece de
fóvea y tiene visión dicromática, el desarrollo, la fisiología, y la estructura general de la retina y
del nervio óptico se parecen mucho a la de los primates, excepto que la lámina cribosa es
menos robusta6, 8, 9.
Es por ello que hemos utilizado el modelo murino para realizar nuestro estudio. De acuerdo
con esto hemos utilizado seis ratones C57BL/6. Además de muestras de ojos humanos
procedentes de la colección del Departamento de Anatomía e Histología Humanas.
Dependiendo de la microscopía empleada, hay diferentes protocolos en relación a las técnicas
específicas que se llevan a cabo:
a) Microscopio óptico (Anexo 1).
b) Microscopio electrónico (Anexo 2)1, 2, 10.
IV. Introducción general de la estructura del globo ocular
Aunque nuestro estudio se centre en el Cuerpo Ciliar consideramos adecuado hacer una breve
introducción a la Anatomía global del globo ocular.
El aparato ocular en su conjunto lo conforman el globo ocular y nervio óptico, y los anexos del
ojo, formados por la musculatura extrínseca y sistema aponeurótico, el aparato lagrimal, la
grasa periocular y los párpados y la conjuntiva.
El globo ocular es un órgano par que tiene forma esférica pero aplanado de arriba abajo,
situado en las cavidades orbitarias. En su parte anterior sobresale un segmento de esfera de
radio menor que el resto del globo ocular que se corresponde con la córnea. Tiene un
diámetro de 23 mm, un peso de 7.5 gr y un volumen de 6.5 cc.
Podemos describir:
Polo anterior: tranparente, corresponde al centro de la córnea.
Polo posterior: localizado en la esclerótica, está lateralmente situado con respecto al
nervio óptico.
Eje del globo ocular: se halla en la línea que une ambos polos.
Ecuador: es el círculo mayor perpendicular al eje. Divide al ojo en dos hemisferios: anterior
y posterior.
Meridianos: círculos que pasan por los dos polos.
Plano antero – posterior: meridiano horizontal. Divide al globo ocular en dos mitades,
temporal y nasal.
4

El globo ocular ocupa la parte anterior de la órbita rebasando todos sus rebordes, excepto el
superior, aunque de forma desigual. Sobresale especialmente por su porción externa, siendo
esta zona la más accesible para la realización de maniobras exploratorias.
Además, el ojo está más próximo a la pared externa que a las otras paredes.
El ojo nunca está en relación directa con los huesos ni con el resto de partes blandas que
contiene la órbita. Se halla recubierto en su parte anterior por los párpados y dorsalmente
alojado en una especie de cápsula fibrosa, la cápsula de Tenon, que lo envuelve y separa del
resto de las estructuras.
El globo ocular está constituido por 3 capas (Fig. 1) de diferente naturaleza que de externa a
interna son: túnica fibrosa, úvea y retina.
Figura 1. Corte semifino. Tinción azul de toluidina, 1. Esclerótica, 2. Úvea, 3. Retina, *. Músculo extrínseco
a) TÚNICA EXTERNA O FIBROSA.
Representa la más superficial de las tres túnicas envolventes del globo ocular. Es una
formación gruesa, resistente y con poca capacidad de extensión. Constituye un mecanismo de
protección para el globo ocular. Comprende dos porciones:
1. Una posterior, opaca: ESCLERÓTICA.
Ocupa los 5/6 posteriores de la túnica fibrosa y es opaca a la luz. Se le puede considerar
como un segmento de esfera hueca, de un color blanco amarillento, que presenta en su parte
anterior una apertura donde se sitúa la córnea y en su parte posterior está atravesada por el
nervio óptico. Tiene un espesor variable, siendo más delgada en su parte media (0.5 mm) pero
más gruesa en las regiones posterior (1 mm) y anterior (0.8 mm).
2. Una anterior, transparente: CÓRNEA.
La córnea es una membrana fibrosa transparente y refringente que se halla encajada en la
abertura anterior de la esclerótica constituyendo 1/6 anterior de la túnica externa. Es un
segmento de esfera hueco cuyo radio menor se adapta a un segmento de esfera de radio
mayor representado por la esclerótica.
En su morfología externa distinguimos una cara anterior, otra posterior y una
circunferencia.
5

Histológicamente formada por seis capas: Epitelio, Membrana de Bowman, Estroma, Capa
de Dua (recientemente descrita, está situada hacia la parte posterior de la córnea, entre el
estroma corneal y la membrana de Descemet. Si bien toda la córnea es de alrededor de 550
micras de espesor, la nueva capa es sólo de alrededor de 15 micras de espesor y se ha
demostrado ser increíblemente fuerte y resistente), Membrana de Descemet y Endotelio.
b) TÚNICA MEDIA O VASCULAR PIGMENTADA, ÚVEA.
Se encuentra adosada a la cara interna de la túnica fibrosa desde el polo posterior hasta 1 mm
del limbo esclerocorneal (coroides). A partir de aquí cambia de dirección situándose
perpendicular al eje del globo ocular (iris), por tanto forma con la córnea un ángulo que recibe
el nombre de ángulo iridocorneal. Esta porción perpendicular marca el límite entre las cámaras
anterior y posterior acabando en un orificio central (pupila).
Distinguimos en la túnica media tres porciones: coroides y cuerpo ciliar, que serán
desarrollados en el apartado de Resultados, e iris.
El iris es la porción más anterior de la úvea, se extiende desde el ángulo de la cámara
anterior hasta el borde pupilar. Está formado por un estroma esponjoso que está en contacto
con la cámara anterior y que constituye la mayor parte del iris, y por una capa epitelial
pigmentada orientada hacia la cámara posterior y que se apoya sobre el cristalino a nivel del
borde pupilar. Además, cerca del borde pupilar, el iris posee un músculo esfinteriano
importante y un tenue músculo dilatador ubicado justo por delante del epitelio pigmentado.
c) TÚNICA INTERNA O NERVIOSA, RETINA.
Es la capa más interna de las tres que constituyen la pared del globo ocular. Su naturaleza es
neural y la consideramos parte del sistema nervioso tanto por su origen como por su función.
Contiene los receptores de la visión siendo la encargada de recoger los estímulos luminosos
que vienen del exterior y transformar los estímulos nerviosos para que sigan su camino hacia
centros visuales encefálicos.
La retina se extiende, concéntrica a la túnica media, desde la salida del nervio óptico hasta la
cara posterior del iris, aunque en el límite entre cuerpo ciliar y coroides pierde sus funciones
neurales.
Distinguimos dos partes en la retina:
1. Retina no neural: se extiende desde la ora serrata hasta la pupila, recubriendo la cara
posterior del cuerpo ciliar y del iris. Esta porción es un resto embrionario de la retina
primitiva que durante el desarrollo ha perdido sus neuronas quedando formada a
expensas de un epitelio pigmentario y otro no pigmentario que estudiaremos junto al
cuerpo ciliar.
2. Retina neural. Es la que se extiende desde la salida del nervio óptico hasta la ora
serrata. Constituye un epitelio pseudoestratificado similar al del tubo neural, el cual
madura produciendo diferentes tipos de neuronas que adoptan una disposición en
capas. Es en realidad la verdadera retina. La morfología interna de la retina visual
presenta una estructura cortical dispuesta en dos estratos:
6

o Epitelio pigmentario de la retina (EPR). Es un epitelio monoestratificado con un
citoplasma cargado de pigmento melánico. Contacta con los segmentos
externos de los fotorreceptores. Este epitelio se encuentra tanto en la retina
neural como en la no neural.
o Estrato neural. Presente solo en la retina neural, y está formado por 10 capas:
epitelio pigmentario, capa de células fotorreceptoras, capa limitante externa,
capa nuclear externa, capa plexiforme externa, capa nuclear interna, capa
plexiforme interna, capa de las células ganglionares, capa de fibras del nervio
óptico y capa limitante interna.
Describimos dos puntos de especial importancia en la retina: la mácula o fóvea y la papila.
1. La mácula o fóvea es un área de aproximadamente 1.5 mm de diámetro ubicada en el
polo posterior, que constituye la zona de mayor agudeza visual y también la zona
óptima para la formación de la imagen en la que encontramos solo conos.
2. La papila del nervio óptico es una zona de más o menos 1 mm de diámetro donde las
fibras nerviosas abandonan el ojo para formar el nervio óptico. En esta región no
existe retina y corresponde a la “mancha ciega” del campo visual.
En el espacio que circunscriben las tres túnicas del ojo se encuentran situados los medios
sólidos y líquidos del ojo (con propiedades transparentes y refringentes) que los rayos
luminosos han de atravesar para llegar hasta la retina: córnea, cristalino, humor acuoso y
humor vítreo.
Estos medios están dispuestos de tal modo que los rayos de luz que parten de un objeto
iluminado, situado en el infinito, han de reunirse en ellos y formar foco en la retina11-15.
V. RESULTADOS: Aportaciones al estudio ultraestructural de coroides
y cuerpo ciliar
a) COROIDES.
La coroides es aquella porción de la úvea que se extiende hacia atrás a partir de la ora serrata.
Se encuentra inmediatamente por dentro de la esclerótica y comprende una capa de coroides
propiamente dicha, muy vascularizada y variablemente pigmentada y una membrana hialina
denominada membrana de Bruch en estrecha relación con la capa pigmentaria de la retina
(Figs. 2 y 3).
7

Figura 2. M.e. (Microscopía electrónica). Ratón. A) Retina: 1. Epitelio Pigmentario Retiniano, 2. Melanosoma, 3. Núcleo célula epitelial, B) Coroides: 4. Membrana de Bruch, 5. Melanocitos y C) Esclerótica: 6. Colágeno,
7. Fibroblastos.
Figura 3. M.e. Ratón. A) Retina: 1. Epitelio Pigmentario Retiniano, *Segmentos Externos de los fotorreceptores,
B) Coroides: 2. Membrana de Bruch, 3. Capilares, 4. Melanocitos y C) Esclerótica.
8

En sus porciones externas, la coroides propiamente dicha contiene vasos relativamente
grandes, en su mayor parte venas; y en sus porciones más internas, inmediatamente antes de
la membrana de Bruch formada por 5 capas bien definidas, presenta una capa simple de
pequeños capilares que se denomina coriocapilar.
La sangre drena hacia el exterior de la coroides por medio de cuatro venas vorticosas, una en
cada cuadrante posterior, y en menor proporción hacia los vasos ciliares anteriores por medio
del cuerpo ciliar. La irrigación de la coroides proviene, en parte, de las arterias ciliares cortas
que entran cerca del nervio óptico, y en parte de las arterias ciliares anteriores que entran en
el ojo desde los músculos extraoculares. Además, la coroides contiene dos arterias ciliares
largas y dos nervios ciliares largos que transcurren hacia el cuerpo ciliar por el meridiano
horizontal.
El estroma de la coroides contiene células pigmentarias, los melanocitos, cuya abundancia
varía según el individuo11, 12, 15.
Los melanocitos dendríticos y los fibroblastos son abundantes en el estroma coroidal, el cual es
bastante delgado en ratones a diferencia con la anatomía humana16.
La coroides se halla compuesta de 4 capas (Fig. 4), que de fuera a dentro son:
o Lámina fusca.
o Capa de los grandes vasos (arterias y venas ciliares).
o Capa de los capilares o coriocapilar.
o Membrana de Bruch.
Figura 4. M.o. (Microscopía óptica). H.e.
(Hematoxolina – Eosina). Humano. 1. Epitelio
Pigmentario Retiniano, 2. Estroma, 3. Coriocapilar,
4. Capa de vasos, 5. Lámina fusca.
9

La célula endotelial, que reviste el lado retiniano de la coriocapilar es extremadamente
delgada y fenestrada (Fig. 5). Las fenestraciones están provistas de diafragmas membranosos.
El citoplasma del lado esclerótico del capilar es grueso y solamente en este lado de la
coriocapilar se encuentran pericitos. El tipo de estructura indica que existe un activo
transporte de líquido desde la coriocapilar hasta el epitelio pigmentario de la retina.
Figura 5. M.e. Ratón. 1. Epitelio Pigmentario Retiniano, Membrana de Bruch (rectángulo), 2. Capa coriocapilar.
La membrana de Bruch (Fig. 6) es una lámina hialina que se interdigita con la coriocapilar en su
superficie posterior, y que constituye la membrana basal del epitelio pigmentario por su cara
anterior. La membrana basal de la coriocapilar constituye, en forma parcial, el límite externo.
La misma consiste de escasas fibras de colágeno y de poco tejido elástico. Cortes planos
muestran que la elástica posee una configuración estrellada y que forma una lámina parecida a
un tamiz en el centro de la membrana. En la edad avanzada la membrana de Bruch contiene
depósitos de lípido y de calcio.
Figura 6. M.e. Ratón. Membrana de Bruch. En la que podemos distinguir 5 capas: Membrana basal de la
coriocapilar, zona colágena laxa externa, capa media de fibras elástica, zona colágena laxa interna y membrana
basal de las células del Epitelio Pigmentario Retiniano.
10
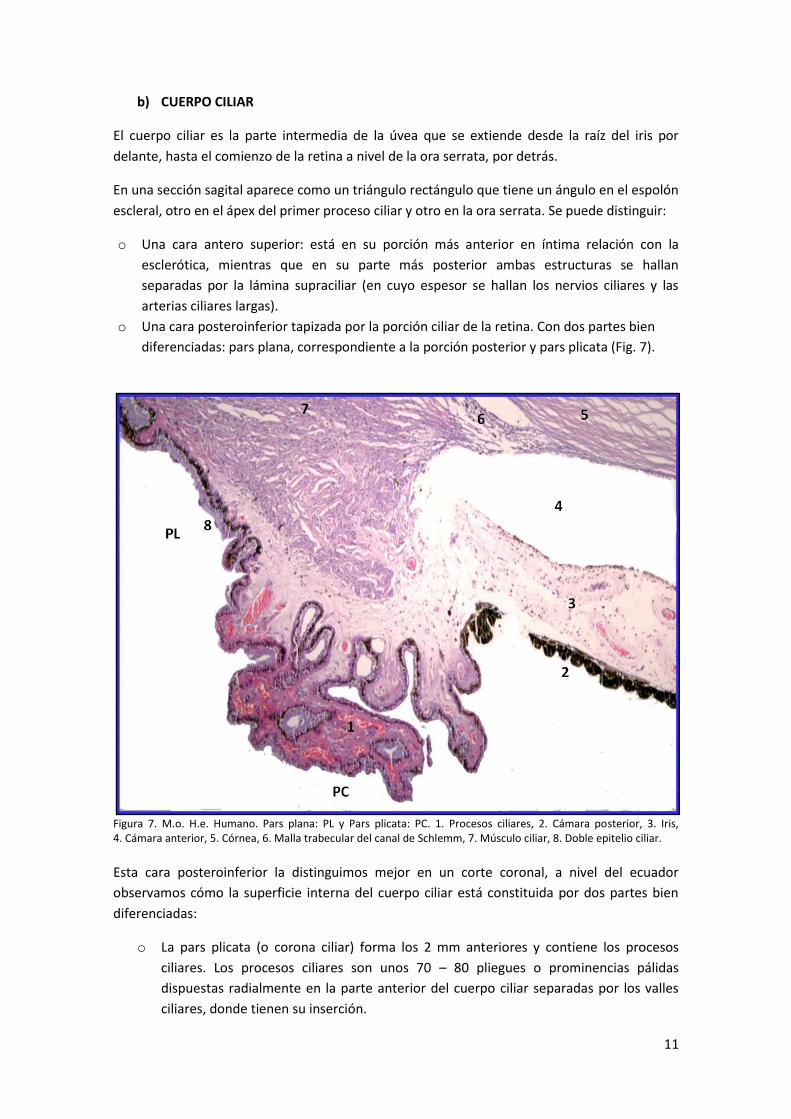
b) CUERPO CILIAR
El cuerpo ciliar es la parte intermedia de la úvea que se extiende desde la raíz del iris por
delante, hasta el comienzo de la retina a nivel de la ora serrata, por detrás.
En una sección sagital aparece como un triángulo rectángulo que tiene un ángulo en el espolón
escleral, otro en el ápex del primer proceso ciliar y otro en la ora serrata. Se puede distinguir:
o Una cara antero superior: está en su porción más anterior en íntima relación con la
esclerótica, mientras que en su parte más posterior ambas estructuras se hallan
separadas por la lámina supraciliar (en cuyo espesor se hallan los nervios ciliares y las
arterias ciliares largas).
o Una cara posteroinferior tapizada por la porción ciliar de la retina. Con dos partes bien
diferenciadas: pars plana, correspondiente a la porción posterior y pars plicata (Fig. 7).
Figura 7. M.o. H.e. Humano. Pars plana: PL y Pars plicata: PC. 1. Procesos ciliares, 2. Cámara posterior, 3. Iris, 4. Cámara anterior, 5. Córnea, 6. Malla trabecular del canal de Schlemm, 7. Músculo ciliar, 8. Doble epitelio ciliar. Esta cara posteroinferior la distinguimos mejor en un corte coronal, a nivel del ecuador
observamos cómo la superficie interna del cuerpo ciliar está constituida por dos partes bien
diferenciadas:
o La pars plicata (o corona ciliar) forma los 2 mm anteriores y contiene los procesos
ciliares. Los procesos ciliares son unos 70 – 80 pliegues o prominencias pálidas
dispuestas radialmente en la parte anterior del cuerpo ciliar separadas por los valles
ciliares, donde tienen su inserción.
11

o La pars plana (o ciliar orbicular) es la parte posterior, más aplanada y de 4 mm de
longitud. Describe unas ondulaciones que terminan formando bahías separadas por
estrías lineales. Está recubierta en su porción más interna por una prolongación
anterior de la retina que a ese nivel sólo consta de dos capas:
La externa pigmentada continuación del Epitelio Pigmentario Retiniano.
La interna no pigmentada que representa el resto de capas neurales de la
retina.
Puede verse que la pars plana se une abruptamente a la retina de una manera
recortada, formando una zona anatómica que denominamos ora serrata (Fig. 8).
Figura 8. Disección del globo ocular, corte coronal. Imagen tomada de Adrian Glasser “disección globo ocular”.
1. Pars Plicata (formada por procesos ciliares), 2. Pars Plana (b. Bahías, el. Estrías lineales), 3. Ora serrata.
El cuerpo ciliar está compuesto por 3 capas: epitelio, estroma vascular y músculo ciliar:
1. Epitelio ciliar
El epitelio ciliar comienza a partir de la ora serrata (punto en el que la retina visual se
transforma en retina ciega) (Fig. 9) y está representado por una doble capa de tejido epitelial
de origen neuroectodérmico, formadas a partir de la zona marginal de la vesícula óptica
embrionaria. La capa más externa se trata de un epitelio pigmentado continuación de la capa
pigmentaria de la retina y que se continúa anteriormente con el epitelio pigmentario del iris,
mientras que la capa interna del epitelio ciliar, cuya membrana basal se apoya sobre el vítreo,
no está pigmentada y se continúa posteriormente con la parte neural de la retina visual (Figs.
10 y 11). Cada capa epitelial tiene una función diferente.
12

Figura 9. M.e. Ratón. Ora serrata. 1. Segmento externo de fotorreceptor, 2. Segmento interno de fotorreceptor,
3. Núcleos de la capa nuclear externa, 4. Ora serrata, 5. Melanocitos, 6. Músculo ciliar, 7. Capilar con célula
endotelial, 8. Glóbulos rojos.
Figura 10. M.e. Ratón. 1. Ora serrata, 2. Epitelio Pigmentario Retiniano, 3. Núcleo de un bastón, 4. Célula bipolar, 5. Segmento interno de fotorreceptor, 6. Segmento externo de fotorreceptor, 7. Epitelio Pigmentario del cuerpo ciliar, 8. Epitelio No Pigmentario del cuerpo ciliar.
13

Figura 11. M.e. Ratón. Ora serrata detalle de la anterior. 1. Núcleos de la capa nuclear externa, 2. Segmento interno de fotorreceptor, 3. Segmento externo de fotorreceptor, 4. Epitelio Pigmentario Retiniano, 5. Músculo ciliar, 6. Melanocitos en la coroides, 7. Transición de Epitelio Pigmentario Retiniano a Epitelio Pigmentario del cuerpo ciliar.
Los extremos apicales de los epitelios pigmentario y no pigmentario están enfrentados.
La base del epitelio ciliar no pigmentado se orienta hacia el vítreo, mientras que la base del
epitelio pigmentario lo hace hacia el estroma vascular. El epitelio ciliar pigmentario tiene una
lámina basal que se fusiona con la lámina basal de los capilares ciliares adyacentes hacia los
que está dirigida, mientras que la lámina basal del epitelio no pigmentario enfrenta la cámara
posterior del globo ocular (Fig. 12).
Figura 12. M.e. Ratón. 1. Capilares fenestrados, 2. Polo basal del Epitelio Pigmentario del cuerpo ciliar, 3. Borde apical del Epitelio Pigmentario, 4. Borde apical del Epitelio No Pigmentario, 5. Polo basal del Epitelio No Pigmentario.
14

La parte basal de las células epiteliales interiores (no pigmentadas) y que enfrenta la
cámara posterior, presenta invaginaciones profundas. El citoplasma de esta zona contiene
numerosas mitocondrias, siendo la parte apical del mismo rica en retículo endoplasmático
granuloso y en aparato de Golgi que refleja un alto nivel de actividad celular. Las vesículas
pinocíticas son también importantes y transportan moléculas de bajo peso molecular. Esta
estructura es coherente con la función de barrera selectiva del epitelio del cuerpo ciliar en la
formación del humor acuoso y la baja presencia de proteínas en su composición. Estas células
además tienen una intensa actividad fagocitótica. Las fibras de la zónula Zinn o ligamento
suspensorio del cristalino se producen fundamentalmente a partir del epitelio ciliar no
pigmentado y se insertan en la membrana basal de estas células, especialmente en los valles
formados por las prolongaciones ciliares y también están unidas a la cápsula de la lente (Fig.
13).
Figura 13. M.e. Ratón. 1. Núcleo, 2. Membrana basal, 3. Microvellosidades de las células epiteliales no pigmentarias,
4. Epitelio Pigmentario, 5. Cromóforo.
Las células pigmentadas de la capa exterior también poseen repliegues basales bien
desarrollados; y en su interior podemos observar abundante presencia de mitocondrias y de
retículo endoplasmático granuloso entre una gran cantidad de gránulos de melanina. (Fig. 14).
En el tejido conectivo entre estos repliegues se encuentran fibras nerviosas. El citoplasma
contiene retículo endoplasmático granuloso (Fig. 15).
15

Figura 14. M.e. Ratón. 1. Capilar fenestrado, 2. Epitelio Pigmentario, 3. Mitocondrias.
Figura 15. M.e. Ratón. A) 1. Melanosomas, 2. Membrana basal del Epitelio Pigmentario, 3. Célula endotelial,
4. Capilar fenestrado, 5. Mitocondrias. B) Detalle de la anterior: 6. Retículo Endoplasmático Granuloso.
El espacio extracelular apical entre las capas del epitelio pigmentario y el epitelio no
pigmentario es mucho más complejo, reflejando las funciones del epitelio ciliar en la
producción del humor acuoso. La mayor parte del espacio está cerrado por uniones
adherentes (zonula adherens) aunque encontramos a menudo pequeñas uniones estrechas
(zonula occludens) entre células adyacentes cerca del borde apical de la célula y también se
pueden observar uniones comunicantes tipo gap (Fig. 16). Los complejos de unión funcionan
como una barrera hemato – acuosa. Sin embargo, en el espacio extracelular a menudo se
forman pequeños espacios intercelulares en parte llenas con vellosidades de procesos
citoplasmáticos, conocidos como los canales ciliares.
16

Figura 16. M.e. Ratón. 1. Zonula adherens, 2. Zonula occludens, 3. Unión gap. En recuadro inferior: Detalle de 3: Unión gap tipo hendidura.
Funcionalmente, el cuerpo ciliar además de producir el humor acuoso y algún
componente del vítreo, es la fuente de las zónulas que soportan la lente. Por lo que está
implicado tanto en el mantenimiento de la presión intraocular como en el proceso de
acomodación (Fig.17).
Figura 17. M.e. Ratón. Epitelio No Pigmentario del cuerpo ciliar. Fibra de zónula de Zinn.
17

2. Estroma vascular
La porción anterior del cuerpo ciliar contiene una arteria circular, el anillo mayor del
iris, que proporciona la irrigación arterial más importante al iris y al cuerpo ciliar. A su vez, se
conecta con las arterias ciliares anteriores y con las arterias ciliares posteriores largas. De este
anillo surgen unas arterias recurrentes que vascularizan todo el cuerpo ciliar capilarizándose a
nivel de los procesos ciliares (Fig. 18).
Figura 18. M.o. H.e. Humano. A) Ángulo iridocorneal. Anillo arterial mayor del iris. B) Detalle de proceso ciliar
capilarizado.
Los capilares que descansan en contacto con el epitelio pigmentario son fenestrados y
permiten un transporte rápido de materiales dentro y fuera de las células del epitelio
pigmentario adyacente así como el humor acuoso formándose un ultrafiltrado selectivo. Existe
un soporte de tejido conectivo que separa el lecho vascular del músculo ciliar y que contiene
abundantes melanocitos dendríticos (Figs. 19 y 20).
Figura 19. M.e. Ratón. Proceso ciliar: Pars plicata. 1. Epitelio No Pigmentado, 2. Epitelio Pigmentado,
3. Capilares.
18

Figura 20. M.e. Ratón. 1. Célula endotelial, 2. Capilar fenestrado, 3. Núcleo célula Epitelial Pigmentada, 4. Nucleolo,
5. Membrana basal y membrana apical del Epitelio Pigmentario.
La estructura del epitelio no pigmentado de la pars plana difiere ligeramente de la pars
plicata. Sus células son altas y sus extremos basales se extienden a menudo hasta el vítreo. Los
repliegues basales son muy amplios y se forman extensos espacios intercelulares. El citoplasma
contiene muchas mitocondrias de gran tamaño, pero sólo una moderada cantidad de
microorganoides membranosos (Fig. 21).
3. Músculo ciliar En un corte transversal, el cuerpo ciliar presenta una forma aproximadamente
triangular, y está ocupado fundamentalmente por una masa de tejido muscular liso, el
músculo ciliar, que controla el enfoque del cristalino. El músculo se subdivide en una banda
circular, situada en el ángulo anterior interno del triángulo, y una porción radial – meridional
que se extiende desde la inserción en la esclerótica, inmediatamente por detrás de la red
trabecular, hasta las porciones posterior e interior del cuerpo ciliar. Las fibras circulares, que se
denominan músculo de Rouger – Müller, relajan la tensión del cristalino y lo acomodan para la
Figura 21. M.e. Ratón. Célula alargada de Epitelio ciliar
No Pigmentario zona posterior.
19

visión cercana; están inervadas por el sistema parasimpático a través del ganglio ciliar. Las
fibras radiales y meridionales (a veces denominadas músculo de Brücke) no tienen una función
claramente demostrada; algunas evidencias sugieren que están inervadas por el sistema
simpático, y que enfocan al cristalino para la visión lejana.
En ratones, el músculo ciliar (Fig. 22) es pequeño y tiene una forma cilíndrica. El
músculo más pequeño está en concordancia con la falta de acomodación en ratones. Las
primeras fibras lisas del músculo pueden ser identificadas justamente anteriores a la
terminación de la retina visual y normalmente se extienden hacia delante justamente al punto
anterior de la terminación posterior del canal de Schlemm. El músculo se demarca
internamente por la base del capilar y externamente por la esclera. Las fibras del músculo
pueden rodear el final posterior del canal de Schlemm14-17.
Figura 22. M.e. Ratón. A, B y C) 1. Fibras musculares, 2. Núcleo, 3. Melanocitos, 4. Capilar, 5. Epitelio No Pigmentario
del cuerpo ciliar, 6. Epitelio Pigmentario del cuerpo ciliar, 7. Lisosomas, 8. Lámina supraciliar.
20

VI. Conclusiones
Como bien he explicado anteriormente, la Anatomía e Histología Ocular es una disciplina
imprescindible para la identificación de las características anatomohistológicas de normalidad
y salud del sistema visual.
Estas dos disciplinas, Anatomía e Histología, son altamente descriptivas y la bibliografía en
ambos campos es extensa, pero se reduce mucho cuando la acotamos al campo del sistema
visual.
No es fácil encontrar documentación sobre imágenes de microscopía electrónica descriptiva de
la calidad de ojos en condiciones de salud, sino que lo que hay es relacionado a las patologías
oculares.
Por lo tanto, lo novedoso que aportaremos en este trabajo serán imágenes de microscopía
electrónica, dada la ausencia de estas como he mencionado anteriormente, así como una
bibliografía en la que podamos observar imágenes de microscopía electrónica descriptiva de la
calidad de ojos en condiciones de salud.
Es por ello que mediante el procesamiento de muestras, tanto para microscopía óptica
convencional como para microscopía electrónica, técnicas histológicas a las que he sido
introducida, he podido revisar la bibliografía existente así como aportar imágenes inéditas
sobre la túnica media o vascular pigmentada, la úvea y, en concreto, sobre la coroides y el
cuerpo ciliar.
Debido a la limitación de extensión del trabajo no he podido abarcar más áreas del sistema
visual como el iris; para abarcar toda la túnica media o vascular pigmentada, ya que de la úvea
disponemos de un gran abanico de imágenes.
21

VII. Bibliografía
1. García del Moral R. Laboratorio de Anatomía Patológica. Madrid: Interamericana
Mc Graw - Hill, 1993.
2. Prieto S, Amich S, Salve ML. Laboratorio clínico. Principios generales. Madrid:
Interamericana Mc Graw - Hill, 1993.
3. Martín N, Martín U. El microscopio [en línea]. [Consulta: 20 Noviembre 2014]. Disponible
en:
http://colegiocristorey.com/nenuca/uca/biologia/biologia_2_bcc/biologia_2_bcc_apunte
s_microscopio.pdf
4. Narváez DJ. La microscopía: herramienta para estudiar células y tejidos [en línea].
[Consulta: 20 Noviembre 2014]. Disponible en:
http://www.medic.ula.ve/histologia/anexos/microscopweb/MONOWEB/capitulo5_2.htm
5. Pei YF, Rhodin JAG. The prenatal development of the mouse eye. Department of
Anatomy, New York Medical College, New York City, 1970.
6. Mackay S, Ballingall CL, Smith RA. P15 Functional characterization of mouse pre – Sertoli
cells cultured on reconstituted basement membrane. Journal of Anatomy.
2002;201(5):430.
7. Tripathi HL, Tripathi N, Kannan GM, Pant BP, Jaiswal DK, Malhotra PR, Flora SJS. Arsenic –
induced changes in certain neurotransmitter levels and their recoveries following chelation
in rat whole brain. Division of Phramacology and Toxicology, Defence Research and
Development Establishment. 1997;92(3):201-208.
8. May CA, Lütjen-Drecoll E. Morphology of the murine optic nerve. Invest Ophthalmol Vis
Sci. 2002;43(7):2206-12.
9. Luesma MJ, Gherghiceanu M, Popescu LM. Telocytes and stem cells in limbus and uvea of
mouse eye. Journal of Cellular and Molecular Medicine 2013;17(8):1016-1024.
10. Parra P, Contamina P. Histología I. Introducción a la Histología [en línea]. [Consulta:
20 Noviembre 2014]. Disponible en:
http://wzar.unizar.es/acad/histologia/textos/TemasHistologia_I/1_1_Introduccion-
Tecnicas.pdf
11. Robles C, Martínez de la Torre M. Guía teórico – práctica para el estudio ocular y de las
vías visuales. Murcia: Diego Marín Librero Editor S.L, 2001.
12. Piñeiro A. Aparato ocular. España: Cusí, 1992.
13. Luesma MJ, Franco A. Bitácora Anatómica e Histológica ocular. Prensas Universitarias de
Zaragoza, 2013.
14. Kaufman PL, Alm A. Adler. Fisiología del ojo. 10ª ed. Madrid: Mosby Elsevier, 2004.
22

15. Weiss L, Greep RO. Histología. 4ª ed. Barcelona: El Ateneo, 1982.
16. Smith RS, Simon WMJ, Nishina PM, Sundberg JP. Systematic Evaluation of the Mouse Eye:
Anatomy, Pathology, and Biomethods. CRC Press, 2001.
17. Gherghiceanu M, Ceafalan LC, Hinescu ME. Celule în alb – negru aide – mémorie de
microscopie electrónica pentro studenti. Bucarest: Medicala Amaltea, 2012.
23


ANEXO 1. Protocolo de las técnicas específicas en microscopía óptica.
1. Disección y fijación
Comenzaremos con la extracción del globo ocular de los ratones C57BL/6.
A continuación, llevaremos a cabo la fijación tisular, la cual consiste en interrumpir los
procesos de degradación que aparecen tras la muerte celular, tratando de conservar la
arquitectura y composición tisular lo más próxima posible a como se encontraba en el
organismo vivo.
La fijación de la muestra tiene un doble objeto:
a) Que los tejidos se preserven lo más parecido a su estado “in vivo”.
b) Que aumente la dureza de la muestra, lo que facilitará su posterior corte en cortes
delgados.
Los principales agentes fijadores que actúan por el mecanismo de reticularización de las
proteínas son aldehídos o potentes oxidantes.
Como agente fijador, el formaldehído se emplea en una concentración del 4 por 100. Sin
embargo, como se parte de una solución de formalina, esta concentración se obtiene
diluyendo una parte de esta última en nueve de agua, solución salina o tampón, durante 4
horas.
Los fijadores hacen que precipiten las proteínas tisulares.
Para continuar, lavado en solución acuosa durante 5 horas.
2. Inclusión
A continuación, pasaremos al proceso que tiene por objeto “rellenar o infiltrar”
completamente la muestra histológica con el medio que se va a utilizar para la imbibición
del tejido, la cual es la infiltración o inclusión en parafina. El fundamento del proceso
radica en la ocupación completa con este medio de los espacios intra y extracelulares
inicialmente rellenos por el agua intratisular. Para conseguir este efecto, casi siempre es
imprescindible eliminar previamente todos los restos de aclarantes residuales en el tejido.
La finalidad última del proceso es proporcionar a la pieza anatómica homogeneidad y
dureza suficiente para que se puedan obtener secciones finas de calidad.
Las parafinas son sustancias de tipo céreo compuestas por mezclas de hidrocarburos
saturados de cadena larga y que pueden obtenerse con una amplia variación en su punto
de fusión (40° – 70°), en gran medida condicionante de sus diferentes aplicaciones en
histotecnología.
El procedimiento de infiltración se realiza mediante baños sucesivos en parafina fundida.
Estos baños deben ser renovados frecuentemente, pues se cargan de solvente (medio de
aclaramiento), cuya presencia provoca un descenso en el punto de fusión de la parafina,
volviéndola más blanda y menos apropiada para el corte.
24
El proceso de infiltración puede realizarse en forma manual o automática, mediante
procesadores de inclusión. Nosotros utilizaremos el manual.
El propósito que guía cualquier proceso de inclusión es conseguir una impregnación lo más
correcta posible en el menor tiempo, sin que la distorsión tisular sea excesiva. En la técnica
manual de inclusión es posible emplear agentes deshidratantes, diafanizantes y líquidos
intermedios.
Los líquidos diafanizantes como es el Benzoato de Metilo, clarifican los tejidos mientras
que los intermedios, como el tolueno, permiten ir reemplazando intratisularmente los
diferentes agentes deshidratantes de manera progresiva.
En la siguiente tabla reproducimos el protocolo de deshidratación, diafanización e
inclusión.
Baños Tiempo para procesamiento manual
Primer día Alcohol 70% 24 horas
Segundo día Alcohol 70% Alcohol 96%
4 – 5 horas Final de la mañana
Tercer día Alcohol 100%
Benzoato de metilo. 1er baño
3 – 4 horas
Cuarto día
Benzoato de metilo. 2° baño Tolueno
Parafina 1ª Parafina 2ª
Media hora 5 minutos
4 horas Final de la mañana
Quinto día Parafina 3ª
Bloques
Mañana siguiente. 2 horas
Como la parafina es un producto inmiscible con el agua hay que deshidratar la muestra
antes de incluirla en la parafina. La deshidratación de la muestra se consigue haciéndola
pasar por alcoholes de gradación progresivamente creciente.
Luego se realizan dos baños en Benzoato de metilo (agente diafanizador) y uno breve en
tolueno (substancia intermedia). Para luego realizar los diferentes baños en parafina
líquida que se recogen en la tabla anterior.
La parafina, cuyo punto de fusión es en nuestro caso de 57°C se encuentra en la estufa de
manera líquida; para realizar los bloques se utilizan moldes o casetes en los que se
deposita la pieza a cortar y se rellenan con la parafina líquida. Siempre hay que observar la
correcta disposición de la pieza en su interior. La orientación de la misma es crucial para su
posterior corte. Los bloques se dejan enfriar a la temperatura ambiente. Luego se guardan
en la nevera hasta su utilización.
25

3. Corte
El micrótomo es un instrumento mecánico con el que se realizan secciones tisulares de
espesor micrométrico y, por tanto, lo suficientemente delgadas para permitir su posterior
observación microscópica de los bloques confeccionados.
Existen seis tipos fundamentales de micrótomos: micrótomo oscilatorio o de balanceo,
micrótomo de rotación o tipo Minot, micrótomo de deslizamiento, micrótomo de
congelación, criostato o criotomo, y ultramicrótomo.
Todos los micrótomos poseen idénticos principios básicos de funcionamiento: un
portabloques en el que se apoya el material que se va a cortar avanza discontinuamente
sobre una cuchilla gracias a un mecanismo regulable de cremallera. De forma periódica se
hace incidir el bloque sobre la cuchilla, obteniéndose secciones tisulares de espesor
equivalente al seleccionado previamente en el tornillo micrométrico que controla el
mecanismo de avance.
El portacuchillas clásico consiste en un dispositivo de fijación basado en una pinza
orientable espacialmente. Esta pinza permite variar el ángulo de inclinación de la cuchilla,
así como su grado de aproximación al portabloques.
Se denomina “mecanismo de avance” del portabloques sobre la cuchilla al sistema
mecánico o electrónico que proporciona cortes sucesivos de tejido a partir del bloque.
Nosotros emplearemos el micrótomo de deslizamiento.
Existen dos tipos fundamentales de micrótomos de deslizamiento, según que se movilice el
portabloques sobre la cuchilla, permaneciendo ésta fija, o viceversa. En ambos casos, el
movimiento que produce el corte es el avance y retroceso de la porción móvil del
micrótomo sobre unas guías metálicas.
La confección de los cortes en parafina consta de seis maniobras o pasos sucesivos, todos
ellos de gran importancia:
1. Retallado y enfriamiento del bloque.
2. Fijación al portabloques.
3. Orientación del bloque.
4. Orientación de la cuchilla del micrótomo y devastado del bloque.
5. Selección del espesor del corte (5 – 10 μm de espesor).
6. Realización de las secciones.
Una vez depositados los cortes sobre portaobjetos, y antes de iniciar el proceso de
coloración, las preparaciones han de ser secadas meticulosamente para evitar su
desprendimiento. La desecación puede realizarse de forma rápida, colocando los
portaobjetos en estufa a 60 °C durante 10 a 20 minutos o, preferiblemente, dejándolas a
37 °C hasta el día siguiente en las placas calefactoras.
26

4. Tinción
Los diversos componentes tisulares tienen unos índices de refracción tan similares entre sí
que no se podrían distinguir unos de otros si no se usaran colorantes que tiñen unos u
otros componentes según su composición química.
Puesto que los colorantes utilizados son solubles en agua y la parafina que embebe los
cortes es inmiscible con el agua hay que desparafinar (disolver y eliminar la parafina,
poniendo los portas en xilol) y rehidratar los cortes haciendo pasar los portas por alcoholes
de gradación progresivamente inferior de acuerdo al siguiente protocolo:
Baños Tiempo para procesamiento
Xilol 1º 10 minutos
Xilol 2º 5 minutos
Xilol 3º 5 minutos
Alcohol 100° 1º 2 minutos
Alcohol 100° 2º 2 minutos
Alcohol 100° 3º 5 minutos
Alcohol 96 2 minutos
Alcohol 70 1º 5 minutos
Alcohol 70 2º 10 minutos
Agua destilada 10 – 15 minutos
Después de la rehidratación se tiñen los cortes con el colorante o la mezcla de colorantes
adecuados, en nuestro caso con hematoxolina – eosina que nos permite observar las
diferentes estructuras del globo ocular.
La hematoxolina (un colorante básico) tiñe de color azul – violeta las estructuras ácidas
(núcleo, ribosomas, retículo endoplasmático granuloso) y la eosina (un colorante ácido)
tiñe de color rosa – rojo las estructuras básicas (la mayoría de las proteínas
citoplasmáticas, mitocondrias). En general, con esta tinción se ven los núcleos de las
células en azul y el citoplasma en rosa. Las fibras de colágena de la matriz extracelular se
tiñen de rosa.
Después de teñir los cortes se aclara el exceso de colorante que quede sin fijar en los
tejidos y se vuelven a deshidratar los cortes.
27

De acuerdo al siguiente protocolo:
Baños Tiempo para procesamiento
Hematoxolina 7 – 8 minutos
Agua corriente 20 minutos – 1 hora
Eosina 30 segundos
Alcohol 70° 1 minuto
Alcohol 96° 1 minuto
Alcohol 100° 1º 1 minuto
Alcohol 100° 2º 2 minutos
Alcohol 100° 3º 2 minutos
Xilol 1º 2 minutos
Xilol 2º 2 minutos
Xilol 3º 5 minutos
Antes de observar los cortes al microscopio hay que cubrirlos con un cubreobjetos de
cristal. Para conseguir que todos los elementos (portaobjetos – preparación –
cubreobjetos) sean ópticamente homogéneos hay que cubrir el corte con un pegamento
(DPX, Permount…), antes de poner el cubreobjetos procurar que no queden burbujas de
aire al colocar éste.
Una vez que el pegamento se ha secado ya se puede examinar la preparación con el
microscopio.
28

ANEXO 2. Protocolo de las técnicas específicas en microscopía electrónica.
1. Fijación
En microscopía electrónica, donde se obtienen imágenes que se cifran en varias decenas
de miles de aumentos, es absolutamente imprescindible, por una parte, la conservación de
las células en el estado más próximo al vivo y, por otra, la obtención de cortes
extremadamente finos (su grosor se mide en decenas de nanómetros) que permiten el
paso del haz de electrones que proyectará la imagen.
Para llevar a cabo la fijación de la muestra, los pequeños bloques de tejido se han
sumergido en una solución tamponada de glutaraldehído al 2.5% porque el glutaraldehído
preserva mejor la estructura de los componentes tisulares. En muchas ocasiones la
solución fijadora se perfunde por vía vascular al animal de experimentación vivo y
anestesiado porque así se consigue una fijación mejor al llegar el fijador a los tejidos
todavía vivos.
Después de lavar los bloques con tampón, se ha realizado una postfijación en tetróxido de
osmio que, además de fijar los tejidos, contrasta las estructuras tisulares por la apetencia
que tienen los lípidos por el osmio.
Se han lavado en tampón fosfato (PB) 0.1 M, cinco baños de 5 minutos. (Si están en fijador
las muestras).
Tras el último lavado se añadió la solución de osmio 2% en PB 0.1 M. y se mantuvieron 1.5
– 2 horas en oscuridad y temperatura ambiente, agitando de vez en cuando (cada 30
minutos). Cuando la solución de osmio comienza a tener color vino retirar
inmediatamente.
Lavar con agua, 3 baños de 5 minutos en la nevera. No exceder de 15 minutos.
2. Inclusión
Los procesos de inclusión son, a grandes rasgos, idénticos a los descritos para la inclusión
general, pues sus procedimientos obedecen a la misma dinámica. No obstante, la
metodología y los medios empleados deben estar en consonancia con el producto final
que se desea obtener: un corte dispuesto para observación. Por las características del
proceso de observación en microscopía electrónica, el corte debe poseer dos cualidades
fundamentales:
a) Una extraordinaria finura o delgadez que permita el paso de los electrones para
proyectar una imagen.
b) Una presentación idónea de la estructura celular y subcelular objeto de
observación.
29

El principio fundamental del proceso de deshidratación es que ha de ser exhaustiva,
porque la mayor parte de los medios de inclusión utilizados son fuertemente hidrófobos y
la persistencia de restos acuosos hace prácticamente imposible la ulterior sección del
bloque.
Por lo general, el número de baños y los tiempos de permanencia en cada uno de ellos
tenderán a ser más cortos que en la inclusión en parafina, dado que el volumen de las
piezas destinadas al estudio con microscopía electrónica es sensiblemente menor que en
el caso de la microscopía óptica.
Nuestro protocolo de inclusión ha sido:
Deshidratar (en nevera):
o Alcohol de 30°: un baño de 5 minutos.
o Alcohol de 50°: un baño de 5 minutos.
o Alcohol de 70°: un baño de 10 minutos.
Lavar en Acetato de Uranilo 2% en alcohol de 70°. El tiempo de incubación en esta
disolución es de 2 horas y media en la nevera. (Favorece el contraste).
Seguir deshidratando (en nevera):
o Alcohol de 70°: dos baños de 5 minutos.
o Alcohol de 96°: dos baños de 10 minutos.
o Alcohol de 100°: dos baños de 10 minutos.
o Alcohol de 100°: un baño de 10 minutos.
Lavar en Óxido de Propileno a temperatura ambiente, llenando casi todo el vial, 2
baños de 10 minutos. No exceder de 30 minutos.
Transferir con un pincel (o pinzas), las muestras desde la solución de óxido de
propileno a la araldita (en moldes de papel de aluminio elaborados en el
momento). Las muestras han de quedar bien sumergidas en araldita y de modo
inmediato. Se pone un poco en el fondo y tras poner la muestra se rellena con
araldita.
Colocar los moldes con muestras en una placa petri de plástico semitapados y se
deja agitando en el shaker (agitador) durante toda la noche.
Meter las muestras en bloques prefabricados (entre portas, cortes < 200 μm; entre
acetatos, cortes de 200 μm; o en tacos).
Polimerizar la araldita durante tres días en estufa de 70°.
30

3. Corte
La observación mediante microscopia electrónica exige un espesor de corte del orden de
algunas decenas de nanómetros debido a la baja capacidad de penetración del haz
electrónico. Por ello, deben obtenerse cortes de grosor inferior a 1 micra, oscilando las
secciones normales entre 30 y 120 nm.
El bloque de resina con la muestra se ponen en un portabloques, se talla el exceso de
resina que hay alrededor de la muestra (se hace una “pirámide”) y se hacen cortes con un
ultramicrótomo que usa cuchillas de vidrio o de diamante.
Se hacen cortes semifinos (de 0.5 – 2 μm de grosor) que se tiñen con azul de
toluidina (1 mg en 1.000 ml agua destilada) y permiten explorar y controlar la zona
que se quiere estudiar con el microscopio electrónico.
Se hacen cortes ultrafinos (de ≈ 50 nm de grosor y una anchura y longitud de 0.5 –
1 mm) que se recogen en una rejilla de cobre o níquel (≈ 3 mm de diámetro).
El ultramicrótomo es un micrótomo básicamente derivado del tipo Minot, el denominado
criostato o criotomo, para cortes en parafina, aunque dotado de numerosas mejoras
técnicas que permiten efectuar secciones de hasta unas pocas decenas de nanómetros de
espesor a partir de material incluido en plástico.
4. Contraste o tinción
Antes de estudiar los cortes con el microscopio electrónico se “tiñen” (en realidad no se
utiliza ningún colorante) o se contrastan los cortes ultrafinos con una solución tamponada
de citrato de plomo (en ocasiones se hace en este momento el contraste con acetato de
uranilo)1, 2, 10.
31








