

CCY Centro de Investigación Científica de Yucatán, A.C.
Posgrado en Ciencias Biológicas
IDENTIFICACIÓN DE LOS miARNs PRESENTES EN
EL DESARROLLO DE LA EMBRIOGÉNESIS
SOMÁTICA DE Coffea canephora.
Tesis que presenta
SARA HERNÁNDEZ CASTELLANO
En opción al título de
MAESTRO EN CIENCIAS
(Ciencias Biológicas: Opción Biotecnología)
Mérida, Yucatán, México
.2014


CENTRO DE INVESTIGACIÓN CIENT{FICA DE YUCA TAN, A. C.
POSGRADO EN CIENCIAS BIOLÓGICAS
CICY RECONOCIMIENTO
POSGRAOOE
CIIE:NCIAS BIOLÓGICAS
Por medio de la presente, hago constar que el trabajo de tesis titulado IDENTIFICACIÓN
DE LOS miARNs PRESENTES EN EL DESARROLLO DE LA EMBRIOGÉNESIS
SOMÁTICA DE Coffea canephora, fue realizado en los laboratorios de la Unidad de
Biotecnología y La Unidad de Bioquímica y Biología Molecular de Plantas del Centro de
Investigación Científica de Yucatán , A.C., bajo la dirección de la Dra. Clelia De la Peña
Seaman, dentro de la opción de Biotecnología de plantas, perteneciente al Programa de
Posgrado en Ciencias Biológicas de este Centro.
Dr. Felipe Augusto Vázquez Flota
Director de Docencia
Mérida, Yucatán, México, Junio 2014.


DECLARACIÓN DE PROPIEDAD
Declaro que la información contenida en la sección de Materiales y Métodos
Experimentales, los Resultados y Discusión de este documento proviene de las
actividades de experimentación realizadas durante el período que se me asignó para
desarrollar mi trabajo de tesis, en las Unidades y Laboratorios del Centro de Investigación
Científica de Yucatán, A.C., y que a razón de lo anterior y en contraprestación de los
servicios educativos o de apoyo que me fueron brindados, dicha información, en términos
de la Ley Federal del Derecho de Autor y la Ley de la Propiedad Industrial, le pertenece
patrimonialmente a dicho Centro de Investigación. Por otra parte, en virtud de lo ya
manifestado, reconozco que de igual manera los productos intelectuales o desarrollos
tecnológicos que deriven o pudieran derivar de lo correspondiente a dicha información, le
pertenecen patrimonialmente al Centro de Investigación Científica de Yucatán, A.C., y en
el mismo tenor, reconozco que si derivaren de este trabajo productos intelectuales o
desarrollos tecnológicos, en lo especial , estos se regirán en todo caso por lo dispuesto por
la Ley Federal del Derecho de Autor y la Ley de la Propiedad Industrial, en el tenor de lo
expuesto en la presente Declaración.
QFB. Sara Hernández Castellano


AGRADECIMIENTOS
Quiero expresar especialmente mi agradecimiento a Dios, a mi familia, amigos y amigas que me han apoyado en las buenas y en las malas, así como a todas las personas que prestaron su colaboración y tiempo durante el desarrollo de este trabajo de investigación.
Al Consejo Nacional de Ciencia y Tecnología, por la beca otorgada No. 334675 para la realización de mis estudios de maestría, y el financiamiento otorgado a través del proyecto de ciencia básica CONACYT-CB2012-178149.
Al Centro de investigación Científica de Yucatán por las instalaciones prestadas, en especial a la unidad de Biotecnología y la Unidad de Bioquímica y Biología Molecular de Plantas por haber proporcionado los equipos y espacios para la realización de este trabajo.
A mi asesora de tesis la Dra. Clelia De la Peña Seaman, por haberme dado la oportunidad de desarrollar este trabajo, su dirección, sus consejos, el tiempo y el apoyo durante la realización de esta tesis.
Al comité tutora! integrado por el Dr. Stefan De Folter, Dr. Enrique Castaño De la Serna y la Dra. Clelia De la Peña Seaman, por sus consejos, observaciones y dirección durante los dos años de trabajo experimental y los cuatro exámenes tutorales.
Al comité evaluador y revisor de tesis integrado por el Dr. Víctor M. Loyola Vargas, El Dr. Stefan De Folter, Dr. Enrique Castaño De la Serna, la Dra. Virginia A. Herrera Valencia y la Dra. Clelia De la Peña Seaman, por sus valiosas observaciones real izadas durante la redacción del documento.
Al Dr. Víctor M. Loyola Vargas por el lugar de trabajo en su laboratorio y el apoyo otorgado para el establecimiento y el mantenimiento del cultivo in vitro de Coffea canephora.
A la Dra. Flor de Fátima Rosas Cárdenas y la M.C. Naivy Gamboa, por el tiempo y las asesorías realizadas en las técnicas de hibridación y marcaje radiactivo .
Al Laboratorio de Epigenética y cromatina de plantas, integrado por el técnico Eduardo Castillo Castro, la M.C. Rosa Y. Us Camas, el I.B.T. Víctor J. Cancino Gárcia, la M.C. Fátima P. Duarte Ake, el Dr. Geovany l. Nic Can y el M.C. Juan Juarez, por su apoyo y comentarios realizados en los seminarios, pero sobre todo, por la amistad que me han brindado durante en estos años.
Al Dr. Geovany l. Nic Can y la M.C. Fátima P. Duarte Ake, por su ayuda técnica, sus consejos y su valiosa amistad que me han brindado.
A mis amigos de seminarios extra oficiales, a mis compañeras (os) de generación 2012-1 y amigas (os) , especialmente a Laura Espinoza, Teresita Valencia, lleana Carrillo , Gamaliel ltza, Fátima Duarte, Rosa Us, Rosita Galaz, Marisa Morales, Miguel Uc, Víctor Cancino, Jorge Xool , Jorge Sonora, Gerardo Manuels, Ulises y a mis amigos tabasqueños (as).

·.·.,.

DEDICATORIAS
El presente trabajo de investigación se encuentra dedicado a papá Dios y a mi familia.
A mi padre y a mi madre pues son la base y fortaleza de mi vida, gracias por ayudarme, por creer en mí, por cada consejo y regaño, pero sobre todo gracias por su amistad y el amor que han compartido con nosotros sus hijos, mamá y papá los amo.
Amis hermanas y hermanos, gracias por ser parte de esta alegría y por el apoyo que me han dado aun en la distancia, por ser más que hermanos (as) mis amigos (as).
A mis princesas, mis hermosas sobrinas Guadalupe y Fátima dos bellos motivos que me impulsaron para alcanzar esta meta.
A mi príncipe, mi bello sobrinito Angel Jesus (+) un ser especial que si bien no se encuentra fisicamente junto ami su amor y enseñanza seguirán siempre en mi vida. Mi bello angel te amo.
A mis tias y tios mas cercanos, gracias por sus palabras de motivación y el cariño que me brindan siempre.
Finalmente le agradezco a Dios por cada prueba, cada lección , cada alegría, cada tristeza que fueron parte de una gran enseñanza que he pasado en estos años de preparación acádemica y personal.
"El hombre encuentra a Dios detrás de cada puerta que la ciencia logra abrir"
Albert Eintein

._,~ .·
. ~ ; •' ...... _ .. .. . -~··..- .. _
.. -. .. ~~.... ;¡; '•of''''
~ '<'i-' ~. '"":~ .. -. '
' ' .l'~ ¡ .. '.\''t,};_:, ... ·¡ d ·; .~ '
;_' . . . ..~., ,~:~
-, .. ;~-;:
·;.('. ",. '. ~ -~~t.-_ .,! .• ., '~- ~-\. .. . (\ ::

INDICE GENERAL
INDICE GENERAL
RESUMEN .... .. . . ... . ........ .. ......... .. . ......... .. ... .. .. ... .... .. .. ... .. ......... ..... ......... .. .. . . . ...... . ...... .. 1
ABSTRACT ............. .. ....... ....... .. . .. .. .... .. ............................ . ...................... .................. III
INTRODUCCIÓN ............................................................................................................... 1
CAPITULO 1: ANTECEDENTES GENERALES .••....••.......•••••••.••....••.••.•.•.••.•..•••.•.•••••••.•.•••. 3
1.1. LOS ARN PEQUEÑOS NO CODIFICANTES . ..... . . .. .... . . .... . .. . .. . ... . ... ........ ..... ... .... .. . .......... 3
1.1.2 BIOGÉNESIS DE LOS MIARNs EN PLANTAS .. .. .... .... .... .. .... .. ................ .. .. .. ...... .. ......... 5
1.1.3 PARTICIPACIÓN DE LOS MIARNS EN EL DESARROLLO DE LAS PLANTAS ............ .. .... .. .. .. . 7
1.1.4 MIARNS Y DESARROLLO EMBRIOGÉNICO IN VITRO ................ .. .............. .. .. .. ............... 8
1.1.5 EMBRIOGÉNESIS SOMÁTICA .............. .. ........ ...... ...... .... ..................... .... ...... .......... .. 9
1.1.6 EL CAFETO ............... ............ .. ... ..... ..... . ....................... ... ... . .. . .......... .. . . . ...... ...... 11
1.1 . 7 EL CULTIVO IN VITRO DE C. CANEPHORA .... .. .. .......... .. .. .. ...... .. .. ................ .. ............ 14
1.2 PREGUNTAS ............................................................................................................ 15
1.3 HIPÓTESIS ................................................................................................................ 16
1.4 OBJETIVOS .............................................................................................................. 16
1.4.1 GENERAL ......................... .. . . .. ... . .. ............. ......... .. . . . ......... .... .... . ....... .. . ............. 16
1.4.2 ESPEC[FICOS .................. .. . .. ... ... .. ... . .. .................. .. .... ........ .. . ......... . ............... ... 16
1.5 JUSTIFICACIÓN ....................................................................................................... 16

!NO/CE GENERAL
1. 7 REFERENCIAS ......................................................................................................... 18
CAPÍTULO 11 IDENTIFICACIÓN DE miARNs CANDIDATOS EN EL DESARROLLO
EMBRIOGÉNICO DE Coffea canephora ........................................................................ 29
2.1 INTRODUCCIÓN ... ... ... ... . .... . .. ........ ... ...... .. . .. .. ........ .. .. . .. . .. ...... . . . .. .. ...................... . .. 29
2.2 MATERIALES Y MÉTODOS ..................................................................................... 30
2.2.1 CULTIVO IN VITRO DE COFFEA CANEPHORA ...... ... . .. ..... ... .. .. .... ........ ... .......... ..... .... .. 30
2.2.2 INDUCCIÓN DE LA EMBRIOGÉNESIS SOMÁTICA A PARTIR DE EXPLANTES FOLIARES DE
COFFEA CANEPHORA ...... . .... ...... . ..... .... ......... .. .. .. . .......... . .. .. . .... .. . .. .. ......... . ... ... ...... .. . .. 31
2.2 .3 EXTRACCIÓN DE LOS ARNS PEQUEÑOS ........ .. .. ..... .. .... ...... .. ................. .......... ...... 31
2.2.4 ANÁLISIS ELECTROFORÉTICO DE MIARNS .... .... .............. .. ... .. .. ....... .... ... .......... .... .. . 32
2.2.5 ANÁLISIS IN SIL/CO DE SECUENCIAS Y DISEÑO DE SONDAS PARA LOS MIARNS DE COFFEA
CANEPHORA . ....... ... . .. .... ... . .... . . . . .. .. . . .. .. .... .. . ... .. ... . .. . . .. . .................. .. . . ... . . . .. . . . .... .. ..... .. 33
2.3 RESULTADOS .......................................................................................................... 34
2.3.1 INDUCCIÓN, DESARROLLO MORFOLÓGICO Y RENDIMIENTO DE LA ES EN COFFEA
CANEPHORA . .......... . . .. .... .. . .. .. . . ......... . .... . . . .......... . ... ........... . .. . ..... .. . ... . ........ ...... .. ... .. .. 34
2.3.2 AISLAMIENTO DE LOS ARN PEQUEÑOS Y MIARNS EN EL PROCESO EMBRIOGÉNICO DE C.
CANEPHORA .. . .. . ..... . ..... ... ........... . .... .. . .............. . .... . ..... . . ... . ..... .. ..... .. .. ......... . .. . . . .. . .... . 36
2.2.3 BúSQUEDA IN SIL/CO DE LOS MIARNS PRESENTES DURANTE EL PROCESO EMBRIOGÉNICO
DE C. CANEPHORA ... .. ... ... . . .. .. . .. .. . . .. .. ................. .. . .. ...... ... .. .... . ..... . ... . ..... ....... ....... . ... .. 37
2.3 DISCUSIÓN .................................................................................... ........................... 38
2.5 REFERENCIAS ......................................................................................................... 44
CAPITULO 111 ANÁLISIS DE LA EXPRESIÓN DIFERENCIAL DE miARNs DURANTE LA

{NO/CE GENERAL
EMBRIOGÉNESIS SOMÁTICA DE C. canephora ........................................................... 53
3.1 INTRODUCCIÓN ...... . . .... .......... ......... .... . . ........ .. .. ... ...... . . ... .. .. . .... . .. . ..... . . . ... .. .......... . 53
3.2 MATERIALES Y MÉTODOS .. .. .. .. ........ . .... . ... . . . . . . ........ . . .... .. . ... . . . .. .. . .. . . . . . .. ... . .. . . .......... . 55
3.2.1 RECUPERACIÓN Y PURIFICACIÓN DE FRAGMENTOS PEQUEÑOS (17 -25 NT) DE ARN ..... . 55
3.2.2 DESFOSFORILACIÓN A PARTIR DE LOS FRAGMENTOS PURIFICADOS DE C. CANEPHORA .. 55
3.2.3 PREPARACIÓN DE MEMBRANAS PARA EL NORTHERN BLOT REVEERSO (NBR) .. . .. . .. . .... 56
3.2.4 ANÁLISIS POR NBR: MARCAJE RADIACTIVO E HIBRIDACIÓN ........................... . . . . . . . ... . . 56
3.2.5 PREPARACIÓN DE LAS MUESTRAS PARA EL ANÁLISIS DEL NORTHERN BLOT
CONVENCIONAL (NBC) ........ . .. .... . ....... . . . .. . . .. ... . . .. .. ............................ . ........... . .. . .. .. . .... 57
3.2.6 PREPARACIÓN DE LAS MEMBRANAS PARA EL NBC ... . .. . . .. ...... . .. ............ . .. . . .. ..... . ...... 57
3.2.7 MARCAJE RADIACTIVO Y PROCESO DE HIBRIDACIÓN .. .... .. . .. . .. .... . .. ...... . . . ... . .... .. .. . .. . .. 57
3.2.8 REVELADO DE LAS PELfCULAS DEL NBR Y NBC .... ... . .... ........ ... .... ...... ..... . .. .. ...... . .... 58
3.2.9. ANÁLISIS DE LA EXPRESIÓN RELATIVA DE LOS MIAR NS DURANTE LA ES ............. .. ..... 58
3.2.9.1 SINTESIS DEL ADN COMPLEMENTARIO (ADNC) Y REACCIÓN DE POLIADENILACIÓN ... . 58
3.2.9.2 ANÁLISIS DE LA EXPRESIÓN DE LOS MIAR NS POR PCR TIEMPO REAL (QPCR) .. ....... . 59
3.2 .9.3 DETERMINACIÓN DE LOS NIVELES DE EXPRESIÓN DE LOS MIARNs . .. .. . .... . ........ . . ..... 59
3.2.1 0 ANÁLISIS ESTADISTICO .... ..... ...... . . ... ... . .. . ....... ... .. . .... . . ... .. . . ... . ...... .... . .. . . .. ..... . . .. .. 59
3.3 RESULTADOS .............................................................................................. ............ 60
3.3.1 ANÁLISIS DEL PATRÓN DE EXPRESIÓN DE LOS MIARNS CANDIDATOS POR NORTHERN BLOT
REVERSO (NBR) ... ..... .............. .... . . . ..... . . .. . . .. . . .... . ... ........... . . . .. .. . . .. . .. ... .... .......... .. .... . .. 60

fNDICE GENERAL
3 .3.2 DETERMINACIÓN DE LOS PATRONES DE EXPRESIÓN POR RT-QPCR DE 16 MIARNS EN
TEJIDOS EMBRIOG~N ICOS DE C. CANEPHORA ............. . . . ......... . .... .. ......... ................ . ....... 64
3 .3 .3 ANÁLISIS DE PATRONES DE EXPRESIÓN SEMI-CUANTITATIVOS POR NORTHERN BLOT
CONVENCIONAL (NBC) ......... .. ..... .. ... . .... .. . .... . .... ............. .......... .... .... . ....... .......... . ...... 71
3.4 DISCUSIÓN .. ... ...... ... .. ......... ...... . ................................... . ....... . ... ...... ... ...... .. .. ........ 72
3.5 REFERENCIAS .. . ... ... . ........ .. ... .... ... . .. .. ........ ... ... .... . .. ..... .. .. ..... .. ..... ...... .. . .. .. . ... ... ... . 79
CAPÍTULO IV: DISCUSIÓN GENERAL, CONCLUSIONES Y PERSPECTIVAS .... ........ 85
4.1 DISCUSIÓN GENERAL ... ....... .. . .... ...... ..... . .. .. ..... .. .......... .. .. ..... . ....... .. . ......... .. .... . 85
4 .2 CONCLUSIÓN GENERAL .. .......... . ...... . ....... .. .. ............ ..... .. .... .. ............. . .. ... ....... . 89
4 .3 PERSPECTIVAS DEL TRABAJO .... ..... ... . ......... ......... ... .. .. ......... . ..... .. .. .. .... .... .. . ... 90
4 .4 REFERENCIAS ......... .. ........ . .......... ..................... ... ... .. ........................ ....... ........ 91
CAPÍTULO V ANEXO: CURVAS DE CALIBRACIÓN Y DETERMINACIÓN DE LA
EFICIENCIA DEL qPCR TIEMPO REAL. ...... .................................... ............................ .. 95

/NO/CE DE FIGURAS
ÍNDICE DE FIGURAS
FIGURA 1.1 Primera clasificación general de los pequeños ARN ...... . .. ... . ..... .... ..... . ... .. 5
FIGURA 1.2. Biogénesis y mecanismo de acción de los miARNs en plantas ...... ........... 6
FIGURA 1.3 Representación del proceso embriogénico somático ... . ... . ... .. .......... .. .. .. .. 11
FIGURA 2.1 Proceso de la ES en Coffea canephora a partir de explantes foliares in vitro ..... .. .... .. .... .. .... . ... ..... . ...... ....... ............... .. ..... . . .. .. .... .... .. .. . ... ..... .. ........... .. 35
FIGURA 2.2 Aislamiento y cuantificación de los embriones somáticos en C. anephora .... 36
FIGURA 2.3 Perfil de bandeo de las extracciones de los pequeños ARN . . .. ... ...... ...... 37
FIGURA 3.1 Niveles relativos de expresión de miARNs conservados en el sistema embriogénico de C. canephora .. . ...... .... ... .. ... .. .. ...... .. ........ .. .. .. ......... ... .. ..... .. .. ... 68
FIGURA 3.2 Niveles relativos de expresión de miARNs conservados en el sistema embriogénico de C. canephora . ... .... .. . .. .. . .... .. ... .... .... . ... . .. ..... .... ..... . . ... ... ..... .. .. . 69
FIGURA 3.3 Niveles relativos de expresión de miARNs divergentes en el sistema embriogénico de C. canephora . .. . ...... ... . ... .. ..... ..... ....... .. .. .. ... ...... .. ... . ... ..... ........ 70
FIGURA 3.4 Detección de los miARNs utilizando el análisis del northern blot convencional durante la ES de C. canephora . ... . ........ . .. .. .. ..... .. .. . . .... ......... ... ... .. ... ... ...... .. .... . .... . 72
FIGURA 4.1 Modulación epigenética de los miARNs en la embriogénesis somática (ES) de Coffea canephora ..... . .. . .. . .. .... ... ...... ... ... ... ... ... ... ... ... ... .. .. .. ... ... ... ... ... ... .... . . ... .. 88


{NO/CE DE CUADROS
ÍNDICE DE CUADROS
CUADRO 2.1 Lista sondas sintetizadas correspondientes a secuencias maduras de miARNs ... .. . .. . .. .. .... . .. ... ..... ... ... ......... .. ...... .................. ... ..... ... ... .. .. ....... .... ..... ... 33
CUADRO 2.2 miARNs conservados e involucrados en el desarrollo de las plantas y su papel durante el proceso de ES .. . .... ... .. ... .. . ....... .. .... .. ............. .. .. . .. .. ................. 41
CUADRO 2.3 miARNs divergentes en el desarrollo de las plantas y su papel durante el proceso embriogénico ..... . ...... .... .. .... ... .. .......... ...... ........ ......................... .. ... . .. 42
CUADRO 2.4 miARNs divergentes involucrados en el desarrollo de las plantas y extremos ambientales .......... ........... .. ... .......... ...... .............. .......... ..... ......... .... .. ................. .. ..... 43
CUADRO 3.1 Patrones de expresión cual itativos mediante la presencia(+) y ausencia(-) de los miARNs evaluados en el proceso embriogénico de C. canephora ........... ........ .. .. . 63
CUADRO 3.2 Clasificación general de los miARNs candidatos analizados en el proceso embriogénico de C. canephora por NBR ..... .... .. ............ .. ...... .. .. .. .... .. .................. .. 64
CUADRO 3.3 Patrones de expresión cuantitativa (qPCR-tiempo real) mínimas (+) ,
intermedias (++) , abundantes (+++)y ausentes (ND) durante el proceso embriogénico de
Coffea canephora .. ......... . .. . .. . ......... ....... .. .. . ........ . .... ......... .......... ........ . ... . ......... 67

.. ·., .. '·
-;;-',;·
,·.

ANEXO
ANEXOS
ANEXO 1 Estandarización las curvas de calibración 5.8'S y snU6 y determinación
de la eficiencia ......................................................................................... 111


RESUMEN
RESUMEN
La embriogénesis somática (ES) es un proceso biológico por el cual las células somáticas
pueden desdiferenciarse y diferenciarse a través de una reprogramación epigenética en
su genoma, estableciendo de esta manera el medio ambiente genético apropiado para el
desarrollo de embriones somáticos con la capacidad de generar una nueva planta. Los
microARNs (miARNs), considerados como elementos claves durante el desarrollo
vegetativo, han sido identificados como elementos esenciales que regulan la respuesta
temprana de la inducción embriogénica somática, sin embargo, el conocimiento acerca de
su papel en el desarrollo de la ES ha sido poco entendido. En el presente trabajo se
analizó el patrón de expresión de 40 miARNs conservados y divergentes durante el
proceso de la ES Coffea canephora.
En los tejidos analizados durante el proceso embriogénico de C. canephora se
identificaron 16 miRNAs por qPCR, sugiriendo su posible función en la ES. Por ejemplo,
m iR 164, miR535, m iR 157 y miR2119 están relacionados con la primera respuesta
generada en los explantes de hojas expuestas al proceso embriogénico. También se
observó que miR164, miR168, miR535 y miR2119 estaban relacionados con la
diferenciación y formación del estadio globular; miR166, miR168, miR393, miR397,
miR398, miR1524 y miR2119 se observaron durante el estadio de corazón, mientras que
la presencia de miR164, miR159 y miR482 se relacionaban con la transición de corazón a
torpedo, y finalmente los m iR 167 y m iR 157 se asociaron con el desarrollo del estadio
cotiledonar. También se observó que la expresión de miR390, miR171, miR156 y miR535
se mantuvieron constantes en la transición del estadio de corazón al de cotiledón. Los
resultados encontrados en este trabajo ponen de manifiesto la importancia de la posible
participación de estos miARNs durante el proceso de ES en C. canephora y proporciona
las bases para el esclarecimiento de la participación de los miARNs en la totipotencialidad
celular en las plantas.

.::·:
.• .-· '.~. ',.
'·
J'. '~. :
... '(;
·'
f'<: ..
' '1'
-~· '·
··./
'. ' ~
-~
'.,
. '

ABSTRACT
ABSTRACT
Somatic embryogenesis (SE) is a biological process by which the somatic cells can be
differentiated and undifferentiated throughout an epigenetic reprogram of the genome,
establishing an appropriate genetic environment to development somatic embryos with the
capacity to generate a new plant. MicroRNAs (miRNAs), considered as key elements
during the vegetative development, have been identified as essential elements regulating
early responses during somatic embryogenic induction. However, the knowledge about the
role of these small molecules during the SE development is poorly understood. In the
present work, it was analyzed the expression pattern of 40 miRNAs, conserved and
divergent, during the SE process of Coffea canephora.
In the analyzed tissues during the embryogenic process of C. canephora, it was identified
16 miRNAs by qPCR, suggesting their possible role in the SE. For instance, m iR 164,
miR535, miR157 and miR2119 were related to the first response generated in the leaf
explants submitted to the embryogenic process. lt was also observed that miR164,
miR168, miR535 and miR2119 were related with the differentiation and the formation of
the globular stage, while that miR166, miR168, miR393, miR397, miR398, miR1524 and
miR2119 were observed during the heart stage development. The presence of miR164,
miR159 and miR482 were related to the transition of the heart to the torpedo stage while
miR167 y miR157 were associated with the development of the cotyledonar stage. lt was
also found that miR390, miR171 , miR156 and miR535 expression were unchanged from
heart to cotyledonar stage. The results found in this work highlight the importance of a
possible role of these miRNAs during the SE process in C. canephora providing the
foundations to elucidate the involvement of these small molecules in the cellular
totipotency of plants.

~ -:
,_:,

INTRODUCCIÓN
INTRODUCCIÓN
Entre los ARN no codificantes se encuentra los ARN pequeños, los cuales tienen
funciones regulatorias muy importantes (Kutter y Svoboda, 2008; Finnegan y Matzke,
2003). Los tres tipos de ARNs reguladores pequeños, actualmente descritos, son los
microARNs (miARNs), los ARNs de interferencia pequeños (siARNs) y los Piwi ARNs
interactivos (piARNs) . Los miARNs son ARN de cadenas sencillas generados
endógenamente y con un tamaño aproximado de 19 a 25 nucleótidos (nt) de longitud.
Hasta el momento se sabe que los miARNs se unen por complementariedad parcial o
total a los ARN mensajeros (ARNm), impidiendo de esta manera su traducción a proteína
o bien generando su degradación (Bartel, 2004; Reinhart et al., 2002; Rajagopalan et al. ,
2006; Allen et al. , 2005; 2008; Chen , 2005). La determinación del papel que desempeñan
estas moléculas y su clasificación han sido posible por los avances realizados para
comprender su biogénesis, la identificación del tipo de precursor, las regiones en las que
se originan y el tipo de complementariedad que ejercen y que influyen directamente sobre
los genes blancos sobre los que ejercen su función regulatoria (Axtell , 2013) .
Para varios grupos de investigación la identificación de los miARNs y el proceso por el
cual se generan estas moléculas, ha tenido un interés particular en la identificación
funcional de estas moléculas sobre el ciclo de vida de las plantas, el papel que
desempeñan sobre la transición y desarrollo tanto en las etapas juveniles como adultas, el
proceso de germinación de las semillas y durante el desarrollo embriogénico (Chen, 2005;
Nonogaki, 2010; Curaba et al., 2012; Giraldez, 2010; Cuperus et al., 201 1). Algunos de los
miARNs reportados y caracterizados hasta el momento están involucrados en los
procesos de desarrollo vegetal con ciertas funciones conservadas entre las especies
mono y eudicotiledóneas (Bonnet et al. , 2004; Lu et al. , 2008; Rhoades, 2012; Cuperus et
al. , 2011).
Estas pequeñas moléculas se encuentran involucradas en la diferenciación y proliferación
celular así como en los mecanismos de respuestas ante extremos bióticos y abióticos
(Chen et al., 2012a; Rogers y Chen , 2013; Wang et al. , 2004). En los últimos años se han
clasificado aproximadamente 20 miARNs, conservados evolutivamente en varias especies
1

INTRODUCCIÓN
de plantas y con funciones específicas sobre el desarrollo y crecimiento vegetal (Chen et
al. , 2012b; Chen et al. , 2011 ; Chi et al. , 2011 ; Kinoshita et al. , 2012; Liu et al. , 2009; Ni et
al. , 2012; Zhang et al., 2012) . Algunos de los miARNs identificados hasta el momento son
miR160, miR168, miR172, miR390 y miR397, los cuales han sido identificados como
elementos esenciales para regular el desarrollo de nuevos órganos en las plantas
(Nonogaki, 201 O; Shen et al., 2009; Buxdorf et al. , 201 O; Reyes y Chua, 2007) . No
obstante, otros miARNs encontrados en procesos como la morfogénesis, la identidad de
órganos florales y el desarrollo embriogénico cigótico se encuentran actuando de manera
tejido-específica (Bonnet et al. , 2004; Mallan na y Rizzino, 201 O; Willmann et al. , 2011) .
El proceso de formación del embrión cigótico, que atraviesa por diversos procesos
bioquímicos y moleculares para el desarrollo de una plántula completa, se encuentra
relacionado al desarrollo embriogénico somático in vitro o embriogénesis somática (ES), y
ha sido util izado como una herramienta efectiva de propagación masiva in vitro de
especies de gran interés económico (Zimmerman, 1993; Koltunow y Grossniklaus, 2003).
La ES es un proceso notable en el que las células somáticas de las plantas desarrollan ,
bajo condiciones controladas, un embrión capaz de convertirse en una planta completa y
funcional (Zimmerman, 1993), y en el que los procesos fisiológicos y moleculares que
envuelven su desarrollo es el resultado de la correcta expresión del programa genético,
(Nassuth et al. , 1980; Hatanaka et al. , 1991 ; Yeung , 1995; Nomura y Komamine, 1995;
Kiran y Thorpe, 1995; Staritsky, 1970). Sin embargo, los estudio epigenéticos realizados
durante el desarrollo de los embriones somáticos y cigóticos han evidenciado a los
miARNs como elementos claves que pueden dirigir o alterar la expresión génica,
determinando el desarrollo o inhibición del proceso embriogénico (Heckel et al. , 1999;
Zhang et al. , 2008; Aquea y Arce-Johnson , 2008)
A partir de esta información, en mi tesis de investigación, se evaluó la expresión de
algunos de los miARNs reportados como conservados en varias especies y que ejercen
funciones importantes durante el desarrollo vegetal y la maduración de embriogénica.
También se consideraron miARNs divergentes pero relacionados al cultivo in vitro, así
como miARNs regulados por diversos factores ambientales y que por primera vez son
estudiados durante el desarrollo de la ES
2

CAPITULO 1
CAPITULO 1: ANTECEDENTES GENERALES
1.1. Los ARN pequeños no codificantes
La regulación en la expresión de genes es un proceso vital en todos los organismos vivos,
y necesario para que se lleven a cabo los procesos de diferenciación celular, la
organogénesis, el desarrollo y la apoptosis celular. Los mecanismos de regulación
genética están basados en la traducibilidad y la estabilidad del ARNm, o bien mediante la
estructuración de la cromatina (Croce y Calin , 2005) . Con la identificación de pequeñas
moléculas de interferencia (ARNi) en las plantas, afloraron otras vías de regulación que
inactivan selectivamente genes de una célula, descifrando así el funcionamiento de cada
gen y su participación en la regulación sobre los patrones expresión (Napoli et al. , 1990;
Sen y Blau, 2006).
El mecanismo de acción dirigido por los ARN interferentes (ARNi) y los ARNs pequeños
no codificantes, permite posicionarlos como elementos principales en la regulación de la
expresión génica, pues regulan aproximadamente un 40% de los genes conocidos (Xie et
al., 201 0). La regulación generada por los ARNs pequeños en el núcleo, como en el
citoplasma, se da sobre la transcripción y la traducción de genes blancos (Matzke y
Birchler, 2005); estas pequeñas moléculas pueden intervenir en todo el proceso de
desarrollo de un organismo, por lo cual ha sido necesario la identificación de su
mecanismo de regulación en células eucariotas (Zhang et al. , 2006; Rhoades et al. , 2006;
Cuperus et al. , 2011) .
Hoy en día se acepta la existencia de tres grandes familias de ARNs pequeños no
codificantes, si bien esta clasificación se encuentra sujeta a cambios con los resultados de
los análisis transcriptómicos a gran escala que se publican diariamente. La primera
clasificación de los ARNs pequeños se basó en su mecanismo de síntesis y la función de
la familia proteica Argonauta (AGO), pues estos fragmentos se asocian a esta familia . Sin
embargo, hoy en día la clasificación general comprende tres clases de pequeños ARNs:
los siARN, los miARNs y los piARNs, estos últimos han sido identificados sólo en
mamíferos (Lu et al. , 2005; Llave et al. , 2002; Kutter y Svoboda, 2008) .
3

CAPITULO 1
En la clasificación más reciente de los siARNs y los miARNs en plantas, los ARNs
pequeños se encuentran en una primera clase que detallada su origen biogénico,
determinado que las secuencias de los siARNs provienen de ARN de doble cadena
(dsARN), en tanto que los miARNs pequeños derivan de ARN de una sola cadena, con la
estructura de ARN de horquilla (hpARNs). Este origen biogénico es en principio el punto
clave que determina gran parte de su papel funcional en las plantas (Chen , 2009; Axtell ,
2013; Allen et al., 2005; Bartel , 2004) (Figura 1.1).
En los últimos años se ha establecido que los siARNs y los miARNs son capaces de
generar un cambio epigenético sobre el mantenimiento e integridad del genoma
(Skopelitis et al. , 2012; Lu et al. , 2005 ; Mallory y Vaucheret, 2006) . Por ello se ha sugerido
que el mecanismo de regulación empleado por estas moléculas se asemeja a la acción de
sustancias morfogénicas en los animales; esto condujo a predecir que estas pequeñas
moléculas de ARN podrían basarse en principios similares para generar, estabilizar e
interpretar sus gradientes de expresión. En general , la primera predicción sobre el papel
que desempeñan los miARN, a partir de las estructuras precursoras en horquilla,
requieren de la acción específica de proteínas con actividad de ARNasa tipo 111 : Drosha y
Dicer, generando miARN maduros de 22 nt, seguido de la acción de AGO, y finalmente se
unen por complementariedad , parcial o total , a los ARNm (Zhang et al. , 2006; Zhang et
al. , 2006; Rhoades, 2012).
4

1} V.ía de los siARNs
ARNds de cadena lar
"P>l RISC
1) Vfa de los miARNs
Precurso e ullo ~p. N
miiARI\I{miAIIN . • d'
to
CAPITULO 1
111) Vía de los piARNs
Figura 1.1 Clasificación de los ARN pequeños. El mecanismo de síntesis a través de la secuencias precursoras, la función de la familia proteica Argonauta (AGO) y el Complejo de Silenciamiento Asociado al ARN (RISC) determinan el tamaño y tipo de secuencias maduras .para los siARN , miARNs y los piARNs. Adaptado de Ghildiyal·y Zamore (2009) .
1.1 .2 Biogénesis de los miARNs en plantas
Hoy en día la importancia que involucra el procesamiento de los miARN y la función
estructural de tallo y horquilla como precursor del transcripto primario (pri-miARN)
demostrada, sugiere que durante la biGgénesis los miARNs maduros proceden a partir de
una cadena no lineal , -llamada MIRGene, precursora en el núcleo, seguida de la etapa
realizada en citoplasma por acción del complejo del silenciamiento asicado al ARN (RISC) y
AG01 , que llevan a cabo la degradación o represión del ARNm a través de la
complementariedad al ARNm (Figura 1.2) (Reinhart et al. , 2002; Bartel , 2004; Chen, 2005;
Westholm y Lai , 2011 ; Baumberger y Baulcombe, 2005; Lasse y Gunter, 2007).
El mecanismo general de la biogénesis y desarrollo de los miARNs en plantas da inicio en
el núcleo, a partir de MIRGEN que se transcribe individualmente pero que contiene una
organización policistrónica para la transcripción. Esta secuencia es considerada como un
promotor para la generación del precursor pri-miARNs, identificando que los extremos 3'-
5

CAPITULO 1
poli-adenilado y 5'- caperuza son reconocidos por la enzima ARN Polimerasa 11 en plantas,
siendo procesadas en el núcleo y generando un fragmento pre-miARN (70nt) y otras
estructuras secundarias (Chen et al. , 2010; Griffiths et al. , 2008; Pegtel et al. , 2010;
Cuperus et al. , 2011 ; Lasse y Gunter, 2007). Asi mismo se ha evidenciado que una
estructura estable del pre-miARNs dentro del núcleo es mantenida por enzimas con
dominio-Dawdle como FHA-nuclear y DDL, en tanto que la ARNse 111 y Dicer Like-1(DCL 1)
generan estructuras apareadas de doble cadena que finalmente y mediante la participación
de Hyponastatic (HYL 1) y C2H2_ Zinc finger Protein Serrate (SE) se estabiliza én el núcleo el
pre-miRNAs. Por lo tanto la interacción sinérgica de HYL 1, SE y DCL 1 en el núcleo (DCL 1)
son elementos esenciales y altamente reconocidos para la bio~énesis de los miARNs
(Carthew et al. , 2009; Rhoades, 2012; Cuperus et"~l. , 2011).
'CITOPLASMA
' Represión del ARNm
Degradación del ARNm
Figura 1.2. Biogénesis y mecanisrl)O de acción de los miARNs en plantas . Modificado de Eldem·et al. , (2013) .
6

CAPITULO 1
1.1.3 Participación de los miARNs en el desarrollo de las plantas
Los miARNs son uno de los componentes importantes en la regulación en todo el ciclo de
vida de las plantas. Los estudios realizados durante la última década han establecido un
papel esencial para los miARNs, sobre el control de expresión génica, las modificaciones
en el epigenoma y los patrones de respuesta contra los virus (Gao et al., 2013; Chen ,
2005; Wang et al., 2001). En el reino vegetal, los miARNs se encuentran conservados
evolutivamente, aunque también existe evidencia sobre miARNs divergentes, tanto en las
plantas como los animales (Zhang et al., 2006; Allen et al. , 2004; Rhoades, 2012; Zhang
et al. , 2006). Estudios realizados sobre los miARNs conservados muestran que esta clase
de miARNs contribuyen al crecimiento adecuado de las plantas y al proceso de la
morfogénesis (Axtell , 2008).
Las evidencia sobre la regulación ejercida por los miARNs muestran a la represión en la
traducción, como un modo de acción regulatoria de los miARNs en las plantas, la llevan a
cabo mediante la complementariedad entre las secuencias maduras de los miRNAs y sus
objetivos blanco (Brodersen et al, 2008.; Dugas y Bartel , 2008; Todesco et al. , 2010).
Otros de los puntos claves sobre el papel de los miARNs en el desarrollo de las plantas,
es la gran similitud entre miembros e isoformas de miARNs de una determinada familia
correlacionándolos funcionalmente (Meng et al., 2011 ; Rubio-Somoza y Weigel, 2011). En
otros estudios, realizados durante el desarrollo de las plantas, se ha centrado la atención
sobre la regulación sinérgica de algunos miARNs, ya que se ha determinado que una
autorregulación de miARNs permite el funcionamiento espacio-temporal de los miARNs;
no obstante, se ha hecho evidente que esta función se encuentra programada a través del
procesamiento nuclear de los pri-miARNs, pre-miARNs y la secuencia madura de los
miARNs (Wang et al. , 2001 ; Hobert, 2008).
En estudios enfocados en la autorregulación que existente entre miembros de una misma
familia o de varios miARNs, se ha identificado la relación funcional entre los miR168a/b
presentes en A. thaliana, pues estos miARNs desempeñan un papel fundamental en la
estabilización de los genes AGO, regulando de esta manera las funciones específicas de
otros miARNs. Fenotípicamente, se ha demostrado que una superposición en el balance
de estos miARNs afectan el desarrollo de la planta, produciendo efectos pleiotrópicos en
condiciones normales de crecimiento (Vaucheret et al. , 2006; Chen, 2009; Meyers et al. ,
7

CAPITULO 1
2008). Si bien la regulación ejercida por el conjunto de miARNs está relacionada
funcionalmente con los factores de transcripción y las familias de genes que presentan
dominios Cup-Shaped Cotyledon (CUC1) y NAC (NAM, ATAF1/2 y CUC2) que a su vez
se encuentran regulados por la familia miR164, son importantes para el desarrollo de la
raíz y los brotes axilares (Raman et al., 2008; Kim et al. , 2009). Así mismo, también se ha
hecho evidente que para el desarrollo correcto de la raíz y el correcto funcionamiento del
factor transcripcional SHORT ROOT (SHR) y las proteínas Scarecrow like (SCR), es
necesaria la regu lación ejercida a través de las familias MIR165 y MIR166 (Williams et al.,
2005; Zhou et al. , 2007; Yanagisawa, 2004) . En varias de las revisiones realizadas de la
función de los miARNs sobre el desarrollo de las plantas se han identificado otras familias
de miARNs, que regulan o reprimen fuertemente otros factores transcripcionales que son
necesarios para el desarrollo de las plantas.
1.1.4 miARNs y desarrollo embriogénico in vitro
La propagación masiva de especies vegetales mejoradas en la naturaleza es un proceso
largo, difícil y costoso. Una alternativa es el uso de técnicas biotecnológicas, como la
propagación in vitro, ya sea por morfogénesis o ES (Santana-Buzzy et al. , 2007, Loyola
Vargas et al. , 2008). Cada una de ellas tiene como base la capacidad totipotente de las
células somáticas del explante (Von Arnold et al. , 2002; Karami et al. , 2009; Karami y
Saidi , 2010; Yeung, 1995).
Si bien el cultivo in vitro es una herramienta para el desarrollo y la micropropagación de
especies económicamente importante, también se puede usar como una herramienta de
estudio para comprender y caracterizar el proceso de desarrollo de las plantas, así como
para estudiar los mecanismos biológicos que permiten su desarrollo embriogénico. La
investigación actual de los miARN pequeños se lha dirigido a aquellos que se encuentran
involucrados directamente en la señalización y que participan en la regulación de los
genes que median el desarrollo de una planta completa (Yeung , 1995). En los últimos
años, durante el proceso de desarrollo embriogénico cigótico y somático, se ha
determinado que estas moléculas desempeñan funciones regulatorias esenciales para el
desarrollo de dichas estructuras embriogénicas al igual que en las hojas, tallos y raíces.
También se ha demostrado que podrían propiciar defectos pleitropicos, pues se cree que
podrían estar regulando más de un tercio de la expresión de todas las proteínas (Palatnik
8

CAPITULO 1
et al., 2003; Ng et al., 2012; Mallanna y Rizzino, 2010; Axtell, 2013; Rogers y Chen, 2013).
El papel regulador de los miARNs durante la embriogénesis somática en plantas todavía
no se entiende bien; hay pocos estudios que han evidenciado el papel de los miARNs
durante el desarrollo de la ES. Durante el proceso de desarrollo embriogénico in vitro
participa un conjunto de vías de señalización muy compleja, por lo que una inadecuada
regulación en ellas, puede dar origen a una reprogramación de la información, inhibiendo
por lo tanto el proceso de ES total o parcialmente. Por ejemplo, usando el cultivo in vitro
de Larix leptolepis como modelo experimental se demostró que una expresión diferencial
del miR171 genera una desregulación en miR172, miR159 y miR169, los cuales están
encargados de activar genes y factores de transcripción durante la diferenciación celular
(Zhang et al., 201 O). Estas observaciones sugieren que un miAR N requiere un sistema de
control de la expresión para permitir el desarrollo de la ES. Otra evidencia proviene de la
identificación de los miARNs y los genes blancos en Dimocarpus logan; en este estudio se
demostró la funcionalidad de diecisiete miARNs durante los diferentes estadios de la
inducción embriogénica, la morfogénesis, y la maduración del embrión cotiledonar (Lin y
Lai, 2013), mientras que en las especies Valencia orange, Medicago truncatula, Pinus
teada y Oryza sativa se identificaron diez miARNs conservados y sus blancos durante el
proceso embriogénico. En resumen, se han podido relacionar ciertos miARNs con los
estadios de inducción y expresión por las que atraviesa el proceso de la embriogénesis
somática (Xiao-Meng et al. , 2011; Aquea y Arce-Johnson, 2008; Eyles et al., 2013), así
como en varios eventos que llevan a la diferenciación celular (Rhoades et al., 2006;
Gutierrez et al., 2007; Willmann et al., 2011).
1.1.5 Embriogénesis somática
En las plantas superiores existen distintas vías de reproducción que conducen al
desarrollo de embriones y a la supervivencia de las especies. Uno de los mecanismos de
reproducción es la vía sexual que se lleva a cabo mediante la fecundación de un gameto
femenino por un gameto masculino, a este proceso se le conoce como embriogénesis
cigótica. Bajo ciertas condiciones, la reproducción de algunas especies vegetales puede
llevarse a cabo en ausencia de la fertilización de los gametos como en el caso de
Ka/anchoe spp. (Mogie, 1992; Koltunow y Grossniklaus, 2003). En condiciones de cultivo
in vitro, la ES es inducida para regenerar una planta completa a partir de una, o un
9

CAPITULO 1
conjunto de células con capacidad totipotente, dando origen a células embriogénicas cuyo
desarrollo asemeja, hasta cierto punto, el proceso de desarrollo de un embrión cigótico
(De Smet et al. , 201 O; Von Arnold et al. , 2002; Staritsky, 1970).
Se ha demostrado que la formación de un embrión somático, que no es el resultado de un
proceso de fertilización , el factor determinante para que la embriogénesis se realice se
encuentra contenido totalmente dentro de la célula , pudiendo funcionar en ausencia de
genes producto del medio ambiente materno o de una señal de fertilización (Nomura y
Komamine, 1995). Las células somáticas de las plantas, por lo tanto, contienen toda la
información genética necesaria para la regeneración de un organismo completo y
funcional mediante la reprogramación celular a través de las condiciones de inducción
embriogénica (Von Arnold et al. , 2002; Nomura y Komamine, 1995).
La presencia de auxinas, ya sea antes o durante el proceso de inducción , el genotipo, el
tipo de explante, la composición del medio, factores físicos como la luz y la temperatura
se han identificado como factores claves para que se lleve a cabo la ES (Karami y Saidi ,
201 O; Karami , et al. , 2009). Una visión molecular sobre los cambios que ocurren desde el
inicio de la ES, hasta la regeneración completa de una planta, han llevado a la búsqueda
de factores relacionados con los mecanismos que controlan la ES, ya que el proceso de
transición de las células somáticas a células embriogénicas, conduce a la activación del
ciclo de división celular, la reorganización fisiológica y al establecimiento de patrones de
expresión para generar células diferenciadas (Von Arnold et al. , 2002; Quiroz-Figueroa et
al., 2006b). En general, durante la ES se podrían identificar dos etapas muy importantes:
la inducción y la expresión, en la primera se adquiere la competencia embriogénica y se
da la proliferación celular, mientras que en la etapa de expresión las células despliegan su
competencia diferenciarse en estructuras somáticas (Quiroz-Figueroa et al. , 2006b).
El proceso de desarrollo de los embriones somáticos es muy semejante al proceso por el
cual se desarrolla un embrión cigótico, excepto que los embriones somáticos no
atraviesan por un periodo de dormancia y desarrollo del suspensor, como es el caso en la
embriogénesis cigótica (Zimmerman, 1993). En la ES las células continúan su
proliferación hasta la obtención de callo embriogénico capaz de diferenciarse
posteriormente en estructuras embriogénicas (Yang y Zhang , 2010) .
10

CAPITULO 1
Coazó
Torped
Plán u a
Figura 1.3 Representación del proceso de embriogénesis som.ática. A partir de explantes de una planta, las células somáticas, bajo condiciones controladas del cultivo in vitro, presentan una reprogramación celular en su epigenoma y en su genoma para dar inicio a la expresión de la totipotencialidad celular y al desarrollo de las estructuras embriogénicas (globular, corazón, torpedo y cotiledonar) (Nic-Can, 2013) .
1.1.6 El cafeto
El cafeto (Coffea spp.)_ es un género perteneciente a la familia de las Rubiaceae, cuenta
con más de 103 especies y ocupa el cuarto lugar como la familia más grande entre las
angiospermas (Davis et al., 2006; Noirot et al. , 2003) . Dos de las principales fuentes de
ingresos y divisas para los países cafeticultores son, C. arabica y C. canephora pues
producen aproximadamente el 75% y 25% de todo el café que se comercializa
mundialmente, respectivamente (Carneiro, 1997). En términos de producción de bebidas,
los consumidores de café favorecen a C. arabica por su sabor menos amargo, delicioso
aroma y menor contenido de cafeína, en comparación con la bebida proveniente de C.
canephora. Sin embargo, la comercialización del café soluble que se consume en la
mayor parte del mundo procede de C. canephora (Herrera et al., 2002).
11

CAPITULO 1
El cafeto se cultiva aproximadamente en 80 países alrededor del mundo y cerca del 70%
de la producción proviene de los pequeños agricultores (Vinod et al., 2006). México es
uno de los países productores de café orgánico, reconocido en el mundo por la calidad del
grano de café y además se ha posicionado entre los mejores a nivel internacionalmente.
El cultivo del cafeto en México, durante el año 2012, representó una de las exportaciones
más lucrativas, pues se exportaron cerca de 3,725,000 sacos con un valor de 875.9
millones de dólares, beneficiando a los pequeños agricultores y familias que dependen de
este cultivo (WWW.sagarpa .gob.mx). En la actualidad , el cafeto es uno de los productos
más valiosos en el comercio internacional. El último informe realizado por la Organización
de las Naciones Unidas para la Agricultura y la Alimentación (FAO)
(http://www.fao.org/economic/est/est-commodities/es) , posiciono al cafeto junto con la
cocoa entre los tres primeros cultivares de mayor importancia en el mercado internacional
después del banano y los cítricos.
Como todo organismo vivo vegetal , el cultivo del cafeto tiene un ciclo de vida y un
potencial productivo característico. El desarrollo vegetativo y reproductivo de una
plantación joven de cafeto necesita aproximadamente de cuatro años para la obtención de
una primera cosecha (semilla - semilla) ; no obstante, estas plantas alcanzan su mayor
productividad a los seis u ocho años de edad y son consideradas comercialmente activas
hasta los 25 ó 30 años, decayendo significativamente la producción de este cultivar
después de esta edad (Arcila et al., 2007; De los Santos-Briones y Hernández-Sotomayor,
2006). Por otra parte, este cultivo puede ser afectado por diversas enfermedades y plagas
que reducen su productividad , en este sentido la especie C. arabica resulta ser más
susceptible a numerosas enfermedades y plagas como: la roya producido por el hongo
Hemielia vastatrix, enfermedades en el fruto por la presencia de Colletrotichum coffanum,
la enfermedad de la broca del café producida por Hypothenemus hampei (mejor conocido
como la broca del café) , plagas como Xylotrechus quadripes Chevr (barrenador del tallo) y
ataque por nemátodos (Meloidogyne spp. y Pratylenchus spp.), mientras que el cultivar de
C. canephora es más resistente a estas enfermedades y plagas (Lashermes et al. , 2008).
En este sentido varios cafeticultores han empleado por mucho tiempo técnicas
convencionales como las retrocruzas , con el fin de generar un genotipo de café estable y
con las características de mayor resistencia a estas enfermedades, sin embargo, el
12

CAPITULO 1
periodo de seis ciclos de auto-polinización implica más de 30 años para generar una
variedad de cafeto mejorado (Carneiro, 1997). En la actualidad se sabe que la limitante
para transferir los rasgos genéticos de resistencia presentes en C. canephora a C.
arabica, mediante técnicas convencionales se debe a que C. canephora es una especie
diploide (2n = 2x = 22) y C. arabica es una especie tetraploide (2n = 4x = 44) (Carneiro,
1999; De-Kochko et al., 201 0). En este sentido los estudios biotecnológicos sobre el
cafeto han perseguido, durante las últimas tres décadas, en laboratorios de todo el
mundo, la mejora agronómica este cultivo.
Durante los primeros estudios mediante la cruza tradicional y el análisis molecular a
través de la selección asistida por marcadores, se logró la identificación y propagación de
genotipos de cafeto deseables en el aumento de la calidad y productividad, sin embargo,
el largo periodo para los ciclos de generación, la exigencia de enormes recursos de la
tierra para los agricultores, el alto costo de las pruebas de campo y la falta de precisión en
la estrategia de esta técnica, ha limitado sustanciamente su mejora (Hendre et al., 2008;
De-Kochko et al. , 201 0) . En la última década los esfuerzos realizados mundialmente para
el mejoramiento del cafeto, han generado ligeros avances pese a las limitantes de los
factores genéticos y fisiológ icos de la misma planta, pues como sabemos, la baja
diversidad genética, el nivel de ploidía, la auto-incompatibilidad y la fácil fertilización entre
las especies robustas, limitan el progreso biotecnológico del cafeto (Carneiro, 1999; Davis
et al., 2006).
No obstante, bajo este contexto y con el uso de herramientas moleculares como la
amplificación aleatoria de ADN polimórfico (RAPO) (Orozco-Castillo et al. , 1996), los
fragmentos de restricción de longitud polimórfica (RFLP) (Pal et al. , 2002), el polimorfismo
en la longitud de fragmentos amplificados (AFLP) (Coulibaly et al. , 2003) y las
repeticiones de secuencias simples o microsatélites (SSR) (Yu et al. , 2011; Poncet et al. ,
2004) se ha logrado el establecimiento de algunos recursos genéticos en cafeto,
avanzado significativamente en la identificación de secuencias expresadas (EST) en
cafeto ( Mondego et al. , 2011; Bhat et al. , 2005) .
Otros de los objetivos biotecnológicos con el cafeto ha sido el mejoramiento sobre el
contenido de cafeína, debido a la sensibilidad de algunos consumidores a este alcaloide
(Silvarolla et al. , 2004). Con base en los estudios realizados por la Asociación Nacional
13

CAPITULO 1
del Café, el consumo del café descafeinado ha superado el 1 O% del café comercializado
en el mundo (http://www.ncausa.org/). Sobre esta necesidad, se ha aplicado la tecnología
del ARN de interferencia (ARNi), para la obtención de plantas transgénicas con una tasa
de síntesis de cafeína reducida (Ogita et al. , 2004; Ogita et al., 2003).
En los primeros estudios realizados sobre la biosíntesis de cafeína, a través de estas
pequeñas moléculas, se avanzó favorablemente en la inhibición en la expresión del gen
CaMXMT1, que codifica para una de las enzimas N-metiltransferasas que está
involucrada en la biosíntesis de la cafeína. El resultado, tras el uso de los ARNi en C.
canephora fue una significativa reducción en el conten ido de cafeína, del 70% al 65%, en
tanto que el uso de la misma tecnología en C. arabica la redujo hasta el 85 al 80% (Ogita
et al. , 2004; Ogita et al. , 2003) . En años recientes, se han idientificado ARNs pequeños,
como los siARNs y los miARNs, en C. canephora y C. arabica con funciones claves en la
ruta de biosíntesis de la cafeína, así como en otros procesos biológicos del cafeto,
abriendo una ruta promisoria para el control en la biosíntesis de este alcaloide.
1.1.7 El cultivo in vitro de C. canephora
Por más de cuatro décadas, el interés por el cafeto se ha convertido en el objeto de
estudio para diferentes líneas de investigación a nivel mundial. Una de ellas, aplicada
para mejorar el cafeto a través de la ingeniería genética ha sido el cultivo in vitro, un
sistema de herramientas que ha permitido estudiar diversos aspectos celulares del café,
tales como químicos (síntesis de cafeína) , fisiológicos (órganos y tejidos) , bioquímicos y
biológicos. Los primeros avances realizados en 1970, se basaron en la ES indirecta
inducida a partir de entrenudos jóvenes y tej idos blandos de C. canephora, algunos años
más tarde se obtuvo la ES en C. arabica. Se utilizó la misma vía de ES indirecta
manipulando secciones de hoja jóvenes (Staritsky, 1970; Sondahl et al. , 1979).
El primer reporte sobre la ES en C. canephora fue publicado por Startsky (1970) y en C.
arabica por Sondal y Sharp (1977). Después, se han publicado varios protocolos. Éstos
pueden ser de un paso (Dublin 1981 ; Yasuda et al. 1985) o una serie secuencial de pasos
en diferentes medios de cultivo (Sondahl y Sharp 1977; Dublin 1984; Zamarripa et al.
1991 ). La inducción de la ES se ha alcanzado usando explantes tales como yemas de
ramas plagiotrópicas y ortotrópicas (Staritsky 1970), explantes de hoja (Sondahl y Sharp
14

CAPITULO 1
1977; Quiroz-Figueroa et al. 2002) , integumentos (Lanaud 1981 ), y anteras y perispermo
(Ascanio y Arcía 1987; Sreenath et al. 1995). Sin embargo, el uso de expalntes de tejido
foliar es lo más común, debido al gran número de explantes obtenidos y la disponibilidad
de tejido a lo largo del año. Además, el tiempo requerido para la inducción de la ES a
partir de diferentes especies de cafeto toma entre ocho meses y un año.
El medio de cultivo usado para la inducción de la ES de cafeto contiene una mezcla de
auxinas y citocininas. Sin embargo, la ES puede inducirse solamente con citocininas
(Dublin 1981 ; Yasuda et al., 1985), puesto que las auxinas tienen un efecto inhibitorio
(Hatanaka et al., 1991 ). Además, Hatanaka et al. , (1995) mostraron que el uso de
inhibidores de la síntesis de etileno (Co2+, Ag+) afectan la ES en cafeto, sugiriendo una
posible función regulatoria para el etileno en la ES de esta especie. El AgN03 mejora la
producción de embriones en cinco genotipos de C. canephora, mientras que a dosis
elevadas tiene un efecto negativo (Fuentes et al. , 2000). El ácido salicílico, a bajas
concentraciones, tiene también un efecto positivo en la calidad y cantidad de los embrioes
obtenidos de C. arabica (Quiroz-Figueroa et al. , 2001 ). Otro compuesto que se ha
determinado también mejora la ES tanto de C. canephora como de C. arabica es el
triacontanol (un alcohol primario de 30 carbonos) (Giridhar et al. , 2004).
En C. canephora se ha determinado que el pre-tratamiento de las plantas cultivadas in
vitre, usadas como fuente de los explantes por 14 días en presencia de ácido
naftalenacético (ANA) y kinetina (KIN) es indispensable para la inducción de la ES.
(Quiroz-Figueroa et al., 2006a). A lo largo de las últimas cuatro décadas, el objetivo
principal de este tipo de estudio ha sido el establecimiento de protocolos eficientes para
regenerar masivamente las especies comerciales de cafeto. También se ha determinado
que factores tales como luz, y temperatura, así como el tipo de explante, el estado
fisiológico , y el pH del medio de cultivo pueden modificar la respuesta embriogénica
(Santana-Buzzy et al. , 2007) .
1.2 PREGUNTAS
1.- ¿Cuáles son los microARNs que están presentes en los diferentes estadios de
desarrollo de los embriones somáticos de C. canephora?
15

CAPITULO 1
2.- ¿Cuál es el patrón de expresión diferencial de los miARN encontrados en los
diferentes estadios del desarrollo embrionario?
1.3 HIPÓTESIS
Si los miARNs son moléculas esenciales involucradas en la morfogénesis celular,
entonces deberán acumularse diferencialmente en los diferentes estadios del proceso de
embriogénesis somática de C. canephora.
1.4 OBJETIVOS
1.4.1 General
);;> Evaluar la presencia de miARNs durante el proceso de embriogénesis somática en
C. canephora.
1.4.2 Especificos
);;> Identificar los miARNs presentes en cada uno de los estadios del proceso
embriogénico en C. canephora.
);;> Analizar la abundancia de miARNs especfficos en cada estadio de desarrollo
embriogénico de C. canephora .
1.5 JUSTIFICACIÓN
El proceso de desarrollo embriogénico conlleva eventos de desdiferenciación y
diferenciación celular, así como otros eventos de adaptación para que se lleve a cabo la
germinación de la semilla y el desarrollo de una planta sana y vigorosa (De Smet et al.,
201 O; Zimmerman, 1993). El desarrollo de los embriones cigóticos y somáticos ha sido
objeto de diversos estudios morfológicos, moleculares, bioquímicos y genéticos. Sin
embargo, recientemente este proceso ha sido abordado bajo el enfoque de los
mecanismos epigenéticos, tales como la metilación del ADN, las modificaciones
postraduccionales en las histonas y la participación de los pequeños ARN no codificantes,
entre los cuales se encuentran los miARNs y los siARNs (Wang et al., 2009; Trionnaire y
Twell , 201 O; Nic-Can et al. , 2013).
16

CAPITULO 1
Se ha observado que los miARNs juegan un papel importante en el desarrollo de las
plantas mediante el control de la expresión de genes blancos. Sin embargo, la función de
estas moléculas durante el proceso de desarrollo de la ES aún no se entiende, y si bien
existen algunos reportes sobre la presencia de los miARNs en la ES en especies como, L.
leptolepis, V. orange, D. /ogan y Oriza sativa, no existe ningún reporte en el que se
discutan cuáles miARN, y en qué cantidad, son expresados en los diferentes estadios
embriogénicos. Es por ello que este estudio puede fortalecer la comprensión del proceso
embriogénico.
1.6 Estrategia experimental
Hoja
•:•Análisis morfológico
,. Caracterización del cultivo in viUo
Aislamiento y colecta de Jos ES a partir del dla 56 posterior a la Inducción ------
Mp Glo Cor Tor
·:·Análisis molecular
,.Extracción de los p~:queños ARNs de bajo peso molecular
.I"Norther blot reverso I"Norther blot convencional
Mediante el marcaje radiactivo con y.:np
I"Sínlesi.s de cONA .l"qRT-PCR-Tiempo
real
Cot Clg
-:·Análisis Bioinformátlco
... Búsqueda e Identificación de miARNs candidatos
.1" Análisis en las bases de datos
.~"Diseño de sondas convencionales
17

CAPITULO 1
1.7 REFERENCIAS
Ascanio , E. , y Arcía , M.M. (1987). Haploids from anther culture in Coffea arabica L. lnternational Congress of Plant Tissue Culture, Tropical Species. Bogotá, Colombia, 68
Allen , E., Xie , Z., Gustafson, A. M., Sung, G.H. , Spatafora, J.W., y Carrington , J.C. (2004). Evolution of microRNA genes by inverted duplication of target gene sequences in Arabidopsis thaliana . Natural Genetic 36, 1282-1290.
Allen , E., Xie, Z., Gustafson, A.M ., y Carrington , J.C. (2005). MicroRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 121, 207-221 .
Altman, S., Ambros, V. , y Appasani , K. (2008). MicroRNAs: From basic science to disease Biology. Cambridge University 1 edition.
Aquea, F., y Arce-Johnson , P. (2008) . ldentification of genes expressed during early somatic embryogenesis in Pinus radiata . Plant Physiology and Biochemistry 46, 559-568.
Arcila , J., Farfán , V.F., Moreno, A.M., Salazar, L.F., y Hincapié, G.E. (2007) . Crecimiento y desarrollo de la planta de café. Sistemas de producción de café en Colombia 22-60.
Axtell , M.J. (2008). Evolution of microRNAs and their targets: Are all microRNAs biologically relevant?. Biochimica et Biophysica Acta 1779, 725-734
Axtell, M.J. (2013). Classification and comparison of small RNAs from plants. Annual Review of Plant Biology 64, 137-159.
Bartel , O.P. (2004). MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116, 281 -297.
Baumberger, N., y Baulcombe, D.C. (2005). Arabidopsis ARGONAUTE1 is an RNA slicer that selectively recruits microRNAs and short interfering RNAs. Proceedings of the National Academy of Sciences of the United S tates of Ame rica 102, 11928-11933.
Bhat, P.R. , Krishnakumar, V., Hendre, P., Rajendrakumar, P., Varshney, R.K., y Aggarwal , R.K. (2005). ldentification and characterization of gene-derived EST-SSR markers from robusta coffee variety 'CxR' (an interspecific hybrid of Coffea canephora and Coffea congensis). Molecular Ecology Notes 5, 83.
Bonnet, E. , Wuyts , J., Rouzé, P., y Van de Peer, Y. (2004). Detection of 91 potential conserved plant microRNAs in Arabidopsis thaliana and Oryza sativa identifies important target genes. Proceedings of the National Academy of Sciences of the United StatesofAmerica 101, 11511-11516.
18

CAPITULO 1
Buxdorf, K. , Hendelman, A. , Stav, R. , Lapidot, M., Ori, N. , y Arazi , T. (2010). ldentification and characterization of a novel m iR 159 target not related to MYB in tomato. Planta 232, 1009-1022.
Carneiro, M.F. (1997) . Coffee biotechnology and its application in genetic transformation . Euphytica 96, 167-172.
Carneiro, M.F. (1999) . Advances in coffee biotechnology. AgBiotechNet 1-7.
Carthew, R.W. , y Sontheimer, E.J. (2009). Origins and mechanisms of miRNAs and siRNAs. Cell 136, 642-655.
Chen, H., Li, Z., y Xiong , L. (2012a) . A plant microRNA regulates the adaptation of roots to drought stress. FEBS Letters 586, 17 42-17 4 7.
Chen, L. , Wang, T. , Zhao, M., Tian, Q., y Zhang , W .H. (2012b) . ldentification of aluminumresponsive microRNAs in Medicago truncatula by genome-wide high-throughput sequencing . Planta 235, 375-386.
Chen, X .M. (2009) . Small RNAs and their roles in plant development. Annual Reviews Cell Development Biology 25, 21-44.
Chen, X ., Zhang , Z. , Liu, D., Zhang , K. , Li, A. , y Mao, L. (2010). SQUAMOSA PromoterBinding Protein-Like transcription factors: Star players for plant growth and development. Journal of lntegrative Plant Biology 52, 946-951.
Chen, X . (2005). microRNA biogenesis and function in plants. FEBS Letters 579, 5923-5931 .
Chen, Z.H., Bao, M.L., Sun, Y.Z. , Yang , Y.J ., Xu , X .H., Wang , J.H. , Han, N. , Bian, H.W., y Zhu, M.Y. (2011). Regulation of auxin response by miR393-targeted transport inhibitor response protein 1 is involved in normal development in Arabidopsis. Plant Molecular Biology 77, 619-629.
Chi, X ., Yang, Q., Chen, X ., Wang, J., Pan, L. , Chen, M., Yang , Z., He, Y. , Liang, X., y Yu, S. (2011 ). ldentification and characterization of microRNAs from peanut (Arachis hypogaea L.) by high-throughput sequencing. Plos One 6, e27530.
Coulibaly, 1., Revol, B., Noirot, M. , Poncet, V., Lorieux, M. , Carasco-Lacombe, C., Minier, J. , Dufour, M., y Hamon, P. (2003). AFLP and SSR polymorphism in a Coffea interspecific backcross progeny [(C. heterocalyx X C. canephora) X C. canephora] . Theoretical and Applied Genetics 107, 1148-1155.
Croce, C.M. y Calin , G.A. (2005) . miRNAs, cancer, and stem cell division . Cell 122, 6-7.
Cuperus, J.T., Fahlgren , N., y Carrington, J.C. (2011) . Evolution and functional diversification of MIRNA genes. The Plant Cell 23, 431-442.
Curaba, J. , Spriggs, A. , Taylor, J. , Li, Z., y Helliwell, C. (2012). miRNA regulation in the
19

CAPITULO 1
early development of barley seed . BMC Plant Biology 12, 120.
Davis, A.P., Goverts, R. , Bridson, D., y Stoffelen , P. (2006). An annotated taxonomic conspectus of the genus Coffea (Rubiaceae) . Botanical Journal of the Linnean Society 152, 465-512.
De los Santos-Briones, C., y Hernández Sotomayor, M.T. (2006) . Coffee biotechnology. Brazilian Journal of Plant Physiology 18, 217-227.
De Smet, 1., La u, S. , M ayer, U. , y Jurgens, G. (201 O) . Embryogenesis - the humble beginnings of plant life. The Plant Journal 61 , 959-970.
De Kochko, A. , Andrade, A.C. , Campa, C., Crouzillat, D., Guyot, R., Hamon, P., Ming, R., Mueller, L.A., y Poncet, V. (201 0) . Advances in coffea genomics. Advances in Botanical Research 53, 23-63.
Dublin P (1981) Embryogenése somatique directe sur fragments de feuilles de caféier Arabusta. Café Cacao Thé 25, 237-242
Dublin P (1984) Techniques de reproduction végétative in vitro et amélioration génétique chez les caféiers cultivés. Café Cacao Thé XXVIII 231-244
Dugas, D.V. y Bartel , B. (2008). Sucrose induction of Arabidopsis miR398 represses two Cu/Zn superoxide dismutases. Plant Molecular Biology 67, 403-417.
Eldem, V., Okay, S., y ünver,T. (2013) . Plant microRNAs: new players in functional genomics. Turkish Journal of Agriculture and Forestry 37, 1-21.
Eyles, R.P., Williams, P.H. , Ohms, S.J ., Weiller, G.F. , Ogilvie, H.A. , Djordjevic, M.A. , y lmin , N. (2013). MicroRNA profiling of root tissues and root forming explant cultures in Medicago truncatula . Planta 238, 91-105.
Finnegan , E.J ., y Matzke, M.A. (2003). The small RNA world . Journal of Cell Science 116, 4689-4693.
Fuentes, S., Calheiros, M. , Manetti , J. , Vieira , L. (2000). The effects of silver nitrate and different carbohydrate sources on somatic embryogenesis in Coffea canephora. Plant Cell Tissue and Organ Culture 60, 5-13.
Gao, R. , Wan , Z.Y., yWong , S.M. (2013). Plantgrowth retardation and conserved miRNAs are correlated to hibiscus chlorotic ringspot virus lnfection . Plos One 8, e85476.
Ghildiyal , M. , y Zamore, P.D. (2009) . Small silencing RNAs: an expanding universe. Nature Review Genetic 10, 94-108.
Giraldez, A. J. (201 O) . MicroRNAs, the cell 's Nepenthe: Clearing the past during the maternal-to-zygotic transition and cellular reprogramming . Current Opinion in Genetics and Development 20, 369-375.
20

CAPITULO 1
Griffiths, J.S., Saini, H.K., Van-Dongen, S. , y Enright, A.J. (2008). miRBase: tools for microRNA genomics. Nucleic Acids Research 36, D154-D158.
Gutierrez, L., Van Wuytswinkel, 0., Castelain, M., y Bellini, C. (2007) . Combined networks regulating seed maturation. Trends in Plant Science 12, 294-300.
Hatanaka, T., Arakawa, 0. , Yasuda, T., Uchida, N., y Yamaguchi , T. (1991). Effect of plant growth regulators on somatic embryogenesis in leaf cultures of Coffea canephora. Plant Cell Reports 10, 179-182.
Hatanaka, T., Sawabe, E., Azuma, T., Uchida, N., y Yasuda, T. (1995) . The role of ethylene in somatic embryogenesis from leaf disks of Coffea canephora. Plant Science 107, 199-204
Heckel, T., Werner, K., Sheridan , W.F., Dumas, C., y Rogowsky, P.M. (1999) . Novel phenotypes and developmental arrest in early embryo specific mutants of maize. Planta 210, 1-8.
Hendre, P., Phanindranath, R., Annapurna, V., Lalremruata, A., y Aggarwal, R. (2008). Development of new genomic microsatellite markers from robusta coffee ( Coffea canephora Pierre ex A. Froehner) showing broad cross-species transferability and utility in genetic studies. BMC Plant Biology 8, 51.
Herrera, J., Combes, M., Anthony, F., Charrier, A., y Lashermes, P. (2002). lntrogression into the allotetraploid Coffea (Coffea arabica L.) : segregation and recombination of the C. canephora genome in the tetreploid interspecific hybrid (C. arabica C. canephora) . Theoretical and Applied Genetics 104, 668-661.
Hobert, O. (2008) . Gene regulation by transcription factors and microRNAs. Science 319, 1785-1786.
Karami, 0 ., Aghavaisi, B., y Pour, A.M. (2009). Molecular aspects of somatic-toembryogenictransition in plants. Journal ofChemical Biology 2, 177-190.
Karami, O., y Saidi , A. (2010). The molecular basis for stress-induced acquisition of somatic embryogenesis. Molecular Biology Reports 37, 2493-2507.
Kim, J.H ., Woo, H.R., Kim, J., Lim, P.O., Lee, I.C., Choi , S.H. , Hwang, D. , y Nam, H.G. (2009). Trifurcate feed-forward regulation of élQe-dependent cell death involving miR164 in Arabidopsis. Science 323, 1053-1057.
Kinoshita, N., Wang, H. , Kasahara, H., Liu , J. , MacPherson, C. , Machida, Y., Kamiya, Y., Hannah, M.A., y Chua, N.H. (2012). /AA-Ala resistant3, an evolutionarily conserved target of miR167, mediates Arabidopsis root architecture changes during high osmotic stress. The Plant Cell 24, 3590-3602.
Kiran , K.S., y Thorpe, T.A. (1995). Asexual embryogenesis in vascular plants in nature. In vitro Embryogenesis in Plants, 17-72.
21

CAPITULO 1
Koltunow, A.M., y Grossniklaus, U. (2003). APOMIXIS: A developmental perspective. Annual Review of Plant Biology 54, 547-574.
Kutter, C., y Svoboda, P. (2008). miRNAs, siRNAs, piRNAs Knowns of the unknown. RNA Biology 5, 181-188.
Lanaud, C. (1981) Production of Coffea canephora plantlets by somatic embryogenesis obtained by in vitro culture of ovules. Café Cacao Thé XXV 231-236
Lashermes, P., Carvalho-Andrade, A, y Etienne, H. (2008). Genomics of coffee one of the world's largest traded commodities. In: Moore Paul H. (ed.), Ming Ray (ed.).Genomics of tropical crop plants. Plant Genetics and Genomics: Crops and Models 203-226 .
Lasse, P., y Gunter, M. (2007). Argonaute proteins: Mediators of RNA silencing. Molecular Cell 26, 611-623.
Lin, Y. , y Lai , Z. (2013). Comparative analysis reveals dynamic changes in miRNAs and their targets and expression during somatic embryogenesis in Longan (Dimocarpus longan Lour). Plos One 8, e60337.
Liu, Q., Zhang , Y.C., Wang, C.Y. , Luo, Y.C., Huang, Q.J. , Chen, S.Y., Zhou , H., Qu, L.H., y Chen , Y.Q. (2009) . Expression analysis of phytohormone-regulated microRNAs in rice, implying their regulation roles in plant hormone signaling . FEBS Letters 583, 723-728.
Loyola-Vargas, V.M., De la Peña, C., Galaz-Ávalos, R.M ., y Quiroz-Figueroa, F.R. (2008). Plant tissue culture. Molecular Biomethods Handbook, 2da edition. Edited by: Walker, J.M. and Rapley, R.Human Press, Totowa, N.J.
Llave, C., Kasschau, K.O., Rector, M.A. , y Carrington, J.C. (2002). Endogenous and silencing-associated small RNAs in plants. The Plant Cell14, 1605-1619.
Lu, C., Tej, S. , Luo, S., Haudenschild, C.D. , Meyers, B.C., y Green , P.J. (2005). Elucidation of the small RNA component of the transcriptome. Science 309, 1567-1569.
Lu, Y., Gan, Q. , Chi , X. , y Qin , S. (2008). ldentification and characterization of microRNAs and their targets in grapevine (Vitis vinífera) . Agricultura! Sciences in China 7, 929-943.
Mallan na, S.K., y Rizzino, A. (201 0). Emerging roles of microRNAs in the control of embryonic stem cells and the generation of induced pluripotent stem cells. Developmental Biology 344, 16-25.
Mallory, A.C., y Vaucheret, H. (2006). Functions of microRNAs and related small RNAs in plants. Nature Genetics 38, S31-S36.
Matzke, M.A. , y Birchler, J.A. (2005). RNAi-mediated pathways in the nucleus. Nature
22

CAPITULO 1
Reviews Genetics 6, 24-35.
Meng, Y., Shao, C. , Wang, H., y Chen , M. (2011). The regulatory activities of plant microRNAs: A more dynamic perspective. Plant Physiology 157, 1583-1595.
Meyers, B.C., Axtell, M.J. , Bartel, B. , Bartel, O.P., Baulcombe, D., Bowman, J.L. , Cao, X., Carrington, J.C., Chen , X., Green, P.J., Griffiths-Jones, S., Jacobsen, S.E., Mallory, A.C., Martienssen, R.A., Poethig, R.S., Qi, Y., Vaucheret, H., Voinnet, O. , Watanabe, Y. , Weigel, D., y Zhu, J.K. (2008). Criteria for annotation of plant microRNAs. The Plant Cell 20, 3186-3190.
Mogie, M. (1992). The evolution of asexual reproduction in plants. Editorial Ilustra, Springer.
Mondego, J.M. , Vidal , R.O., Carazzolle, M.F., Tokuda, E.K., Parizzi , L.P., Costa, G.G. , Pereira, L.F., Andrade, A.C., Colombo, C.A., y Vieira, L.G. (2011). An EST-based analysis identifies new genes and reveals distinctive gene expression features of Coffea arabica and Coffea canephora. BMC Plant Biology 11 , 30.
Napoli , C., Lemieux, C. , y Jorgensen, R. (1990). lntroduction of a chimeric chalcone synthase gene into petunia results in reversible co-suppression of homologous genes in trans. The Plant Cell 2, 279-289.
Nassuth, A. , Wormer, T.M. , Bouman, F., y Staritsky, G. (1980) . The histogenesis of callus in Coffea canephora stem explants and the discovery of early embryoid initiation. Acta Botanica Neerlandica 21, 49-54.
Ng, D.W. , Lu, J., y Chen, Z.J. (2012). Big roles for small RNAs in polyploidy, hybrid vigor, and hybrid incompatibility. Current Opinion in Plant Biology 15, 154-161.
Ni, Z. , Hu, Z., Jiang, Q., y Zhang , H. (2012). Overexpression of gma-MIR394a confers tolerance to drought in transgenic Arabidopsis thaliana. Biochemical and Biophysical Research Communications 427, 330-335.
Nic-Can, G.l. (2013). Análisis de los cambios epigenéticos asociados con la diferenciación celular en la embriogénesis somática de Coffea canephora . Universidad Autónoma de Yucatán. Tesis.
Nic-Can, G.l. , López-Torres, A., Barreda-Pool, F., Wrobel , K., Loyola-Vargas, V.M. , RojasHerrera, R., y De-la-Peña, C. (2013). New lnsights into somatic embryogenesis: LEAFY COTYLEDON1, BABY BOOM1 and WUSCHEL-RELA TED HOMEOBOX4 are epigenetically regulated in Coffea canephora. Plos One 8, e72160.
Noirot, M. , Poncet, V. , Barre, P., Hamon, P. , Hamon, S., y de Kochko, A. (2003). Genome size variations in diploid African Coffea species. Annals of Botany 92, 709-714.
Nomura, K. , y Komamine, A. (1995). Physiological and biochemical aspects of somatic embryogenesis. In vitre embryogenesis in plants, T.A.Thorpe, ed. Dordrecht: Kluwer Academic Publishers 249-265.
23

CAPITULO 1
Nonogaki, H. (201 0) . MicroRNA gene regulation cascades during early stages of plant development. Plant and Cell Physiology 51 , 1840-1846.
Ogita, S., Uefuji, H. , Yamaguch i, Y. , Koizumi , N., y Sano, H. (2003). RNA interference: producing decaffeinated coffee plants. Nature 423, 823-836.
Ogita, S., Uefuji, H. , Morimoto, M., y Sano, H. (2004). Application of RNAi to confirm theobromine as the major intermediate for caffeine biosynthesis in coffee plants with potential for construction of decaffeinated varieties. Plant Molecular Biology 54, 931-941.
Orozco-Castillo , C. , Chalmers, K.J ., Powell , W ., y Waugh, R. (1996). RAPO and organelle specific PCR re-affirms taxonomic relationships within the genus Coffea. Plant Cell Reports 15, 337-341 .
Palatnik, J.F, Allen , E. , Xuelin , W. , Schommer, C. , Schwab, R., Carrington , J. , y Weigel , D. (2003). Control of leaf morphogenesis by microRNAs. Nature 425, 257-263 .
Pal , N., Sandhu , J.S., Domier, L.L., y Kolb , F.L. (2002) . Development and characterization of microsatellite and RFLP-derived PCR markers in oat. Crop Science Society of America 42, 912-918.
Pegtel, D.M., Cosmopoulos, K. , Thorley-Lawson, D.A. , Van-Eijndhoven, M.A. , Hopmans, E.S., Lindenberg , J.L. , De Gru ij l, T.D., Würdinger, T. , y Middeldorp, J.M. (2010). Functional delivery of viral miRNAs via exosomes. Proceedings of the National Academy of Sciences 107, 6328-6333.
Poncet, V., Hamon, P., Minier, J., Carasco, C., Hamon, S., y Noirot, M. (2004). SSR crossamplification and variation within coffee trees (Coffea spp.). Genome 47, 1071-1081 .
Quiroz-Figueroa, F.R. , Fuentes-Cerda, C.F., Rojas-Herrera , R. , y Loyola-Vargas, V.M. (2002). Histological studies on the developmental stages and differentiation of two different somatic embryogenesis systems of Coffea arabíca. Plant Cell Reports 20, 1141-1149.
Quiroz-Figueroa, F.R. , Méndez-Zeel, M., Larqué-Saavedra, A. , y Loyola-Vargas, V.M. (2001 ). Picomolar concentrations of salycilates induce cellular growth and enhance somatic embryogenesis in Coffea arabíca tissue culture. Plant Cell Reports 20, 679-684.
Quiroz-Figueroa, F.R., Monfore-Gonzáles, M. , Galaz-Avalos, R.M., y Loyola-Vargas, V.M. (2006a). Direct somatic embryogensis in coffea canephora . Loyola-Vargas, V.M.y Vásquez-Fiota, F.A. eds. Totowa, New Jersey 111-117.
Quiroz-Figueroa, F.R. , Rojas-Herrera, R. , Galaz-Avalos, R.M., y Loyola-Vargas, V.M. (2006b). Embryo production through somatic embryogenesis can be used to study cell differentiation in plants. Plant Cell Tissue and Organ Culture 86, 285-301.
24

CAPITULO 1
Rajagopalan, R. , Vaucheret, H. , Trejo, J., y Bartel , O.P. (2006) . A diverse and evolutionarily fluid set of microRNAs in Arabidopsis thaliana. Genes and Development 20, 3407-3425.
Raman , S., Greb, T. , Peaucelle, A. , Blein, T., Laufs, P. , y Theres, K. (2008). lnterplay of miR164, CUP-SHAPED COTYLEDON genes and LATERAL SUPPRESSOR controls axillary meristem formation in Arabidopsis thaliana . The Plant Journal 55, 65-76.
Reyes, J.L., y Chua, N. (2007). ABA induction of miR159 controls transcript levels of two MYB factors during Arabidopsis seed germination . The Plant Journal 49, 592-606.
Reinhart, B.J., Weinstein , E.G., Rhoades, M.W., Bartel , B., y Bartel, O.P. (2002). microRNAs in plants. Genes and Development 16, 1616-1626.
Rhoades, M.W ., Reinhart, B.J., Lim, L.P., Surge, C.B., Bartel , B., y Bartel , O.P. (2002). Prediction of plant microRNA targets. Cell 11 O, 513-520.
Rhoades, M.W. (2012). Conservation and divergence in plants microRNAs. Plant Molecular Biology 80, 3-16.
Rhoades, J., Bartel, O.P. , y Bartel, B. (2006) . miRNAs and their regulatory roles in plants. Annual Review of Plant Physiology 57, 19-53.
Rogers, K., y Chen, X. (2013). Biogenesis, turnover, and mode of action of plant microRNAs. The Plant Cell 25, 2383-2399.
Rubio-Somoza, 1. , y Weigel , D. (2011). microRNA networks and developmental plasticity in plants. Trends in Science 16, 258-264.
Samson, N.P. , Campa, C. , Gal, L.L., Noirot, M. , Thomas, G., Lokeswari , T.S., y Kochko, A. (2006). Effect of primary culture medium composition on high frequency somatic embryogenesis in different Coffea species. Plant Cell Tissue and Organ Culture 86, 37-45.
Santana-Buzzy, N., Rojas-Herrera , R. , Galaz-Avalos, R.M. , Ku-Cauich , J.R., MijangosCortés, J., Gutiérrez-Pacheco, L.C., Canto, A. , Quiroz-Figueroa, F., y LoyolaVargas, V.M. (2007). Advances in coffee tissue culture and its practica! applications. In Vitro Cellular and Developmental Biology-Piant 43, 507-520.
Sen, G.L. , y Blau , H.M. (2006) . A brief history of RNAi : the silence of the genes. The FASES Journal 20, 1293-1299.
Shen, D. , Wang , S. , Chen, H., Zhu, Q.H. , Helliwell , C., y Fan , L.J. (2009). Molecular phylogeny of miR390-guided trans-acting siRNA genes (TAS3) in the grass family. Plant Systematics and Evolution 283, 125-132.
Silvarolla, M.B. , Mazzafera, P. , y Fazuoli , L.C. (2004). Plant biochemistry: A naturally decaffeinated arabica coffee. Nature 429, 826-836.
25

CAPITULO 1
Skopelitis, D.S. , Husbands, A.Y., y Timmermans, M.C. (2012). Plant small RNAs as morphogens. Current Opinion in Cell Biology 24, 217-224.
Small , l. (2007) . RNAi for revealing and engineering plant gene functions. Current Opin ion in Biotechnology 18, 148-153.
Sondahl, M.R. , Sharp, W.R. (1977) High frequency induction of somatic embryos in cultured leaf explants of Coffea arabica L. Zeitschrift für Pflanzenphysiologie 81, 395-408
Sondahl, M.R. , Salisbury, J.L. , y Sharp, W.R. (1979). SEM characterization of embryogenic tissue and globular embryos during high frequency somatic embryogenesis in coffee callus cells. Zeitschrift für Pflanzenphysiologie 94, 185-188.
Sreenath, H.L. , Shanta, H.M. , Babu, K.H., Naidu, M.M. (1995) Somatic embryogenesis from integument (perisperm) cultures of coffee. Plant Cell Reports 14, 670-673
Staritsky, G. (1970). Embryoid formation in callus tissues of coffea. Acta Botanica Neerlandica 19, 509-514.
Townsley, P. (1974). Production of coffee from plant cell suspension cultures. Journal de l'lnstitut Canadien des Sciences et Technologies d'Aiimentation 79-81 .
Trionnaire, G.L,. y Twell , D. (2010). Small RNAs in angiosperm gamethophytes: from epigenetics to gamete development. Genes and Development 24, 1081-1085.
Van Boxtel, J., y Berthouly, M. (1996) . High frequency somatic embryogenesis from coffee leaves. Plant Cell Tissue and Organ Culture 7-17.
Vaucheret, H., Mallory, A.C. , y Bartel , O.P. (2006). AG01 homeostasis entails coexpression of MIR168 and AG01 and preferential stabilization of miR168 by AG01 . Molecular Cell 22, 129-136.
Vinod, K., Naidu, M. , y Ravishankar, G.A. (2006) . Developments in coffee biotechnology in vitro plant propagation and crop improvents. Plant Cell Tissue and Organ Culture 87, 49-65.
Von Arnold , S., Sabala, 1., Bozhkov, P. , Dyachok, J., y Filonova, L. (2002) . Developmental pathways of somatic embryogenesis. Plant Cell Tissue and Organ Culture 69, 233-249.
Wang , W.X., Gaffney, B., Hunt, A.G. , y Tang , G. (2001). microRNAs (miRNAs) and plant development. In eLS, John Wiley and Sons.
Wang , X. , Elling , A.A. , Li, X. , Li , N., Peng , Z. , He, G., Sun, H., Qi , Y., Liu, X.S. , y Deng, X.W. (2009) . Genome-wide and organ-specific landscapes of epigenetic modifications and their relationships to mRNA and small RNA transcriptomes in Maize. The Plant Cell 21, 1053-1069.
26

CAPITULO 1
Wang, X.J., Reyes, J.L., Chua, N.H., y Gaasterland ,T. (2004). Prediction and identification of Arabidopsis thaliana microRNAs and their mRNA targets. Genome Biology 5, R65.
Wassenegger, M. (2005). The role of the RNAi machinery in heterochromatin formation. Cell122, 13-16.
Westholm, J.O. , y Lai, E.C. (2011). mirtrons: MicroRNA biogenesis via splicing. Biochimie Journal 93, 1897-1904.
Williams, L., Grigg , S.P., Xie, M., Christensen , S., y Fletcher, J.C. (2005). Regulation of Arabidopsis shoot apical meristem and lateral organ formation by microRNA miR166g and its AtHD-ZIP target genes. Development 132, 3657-3668.
Willmann, M.R. , Mehalick, A.J., Packer, R.L., y Jenik, P.D. (2011). microRNAs regulate the timing of embryo maturation in Arabidopsis. Plant Physiology 155, 1871-1884.
Xiao-Meng , Wu., Mei-Ya, L. , Xiao-Xia , Ge., Qiang , Xu ., y Wen-Wu, G. (2011 ). Stage and tissue-specific modulation of ten conserved miRNAs and their targets during somatic embryogenesis of Valencia sweet orange. Planta 233, 495-505.
Xie, Z., Khanna, K., y Ruan, S. (2010). Expression of microRNAs and its regulation in plants. Seminars in Cell and Developmental Biology 21, 790-797.
Yanagisawa, S. (2004) . Dof domain proteins: plant-specific transcription factors associated with diverse phenomena unique to plants. Plant and Cell Physiology 45, 386-391.
Yang, X., y Zhang , X.L. (2010). Regulation of somatic embryogenesis inhigher plants. Critica! Reviews in Plant Sciences 29, 36-57.
Yasuda, T., Fujii, Y., y Yamaguchi, T. (1985). Embryogenic callus induction from Coffea arabica leaf explants by benzyladenine. Plant and Cell Physiology 26, 595-597.
Yasuda, T., Tahara, M., Hatanaka, T., Nishibata, T. , y Yamaguchi, T. (1995). Clona! propagation through somatic embryogenesis of Coffea species. XVI Colloquial Science lnternational Coffee Kyoto 392.
Yeung, E.C. (1995). Structural and development patterns in somatic embryogenesis. In Vitro embryogenesis in plants, T.A. Thorpe, ed . Netherlands: Kluwer Academic Publishers 205-247.
Yu, Q., Guyot, R., Kochko, A., Byers, A., Navajas-Perez, R., Langston, B.J. , DubreuiiTranchant, C., Paterson, AH., Poncet, V., y Nagai, C. (2011). Micro-collinearity and genome evolution in the vicinity of an ethylene receptor gene of cultivated diploid and allotetraploid coffee species (Coffea). The Plant Journal 67, 305-317.
Zamarripa, C.A., Ducos, J.P. , Tessereau, H., Bollon H, Eskes, A.B., Pétiard, V. (1991) Devéloppement d'un procédé de multiplication en masse du caféier par embryogenése somatique en milieu liquide. Anonymous, 14e Colloque Scientifique
27

CAPITULO 1
lnternationale sur le Café, Association Scientifique lnternationale du Café, Paris 392-402
Zhang, 8. , Pan , X ., Cannon, C. H., Cobb, G.P. , y Anderson, T.A. (2006) . Conservation and divergence of plant microRNA genes. The Plant Journal 46, 243-259.
Zhang , 8. , Pan, X. , y Stellwag , E.J. (2008). ldentification of soybean microRNAs and their targets. Planta 229, 161-182.
Zhang, J. , Zhang , S. , Han, S. , Wu, T. , Li, X. , Li , W., y Qi , L. (2012). Genome-wide identification of microRNAs in larch and stage-specific modulation of 11 conserved microRNAs and their targets during somatic embryogenesis. Planta 236, 647-657.
Zhang , S., Zhou, J., Han, S., Yang , W. , Li , W ., Wei , H., Li , X. , y Qi , L. (2010). Four abiotic stress-induced miRNA families differentially regulated in the embryogenic and nonembryogenic callus tissues of Larix /eptolepis. 8iochemical and 8iophysical Research Communications 398, 355-360.
Zhou, G.K. , Kubo, M., Zhong , R. , Demura, T., y Ye, Z.H. (2007). Overexpression of m iR 165 affects apical meristem formation , organ polarity establishment and vascular development in Arabidopsis. Plant and Cell Physiology 48, 391-404.
Zimmerman,J.L. (1993). Somatic embryogenesis: A model for early development in higher plants. The Plant Cell 5, 1411-1423.
28

CAPITULO 11
CAPÍTULO 11 IDENTIFICACIÓN DE miARNs CANDIDATOS EN EL
DESARROLLO EMBRIOGÉNICO DE Coffea Canephora
2.1 Introducción
El desarrollo embriogénico vegetal comprende varios procesos biológicos que permiten
establecer sistemas de señales transcripcionales y generar los patrones funcionales y
estructurales que determinan el correcto desarrollo de la estructura embriogénica
(Kapinas et al., 2013; Cangahuala-lnocente et al., 2009). La capacidad de producir un
organismo completo, a partir de células somáticas indiferenciadas, reside únicamente en
el reino vegetal (Zimmerman, 1993; Quiroz-Figueroa et al. , 2006b) . En la naturaleza, y
bajo ciertas condiciones, se puede llevar a cabo el desarrollo de embriones en ausencia
de fertilización, este proceso recibe el nombre de apomixis. En algunas especies los
embriones surgen de las hojas como propágulos vegetativos (p. ej. Ka/anchoe spp.). Los
embriones surgen directamente de una célula somática. En condiciones in vitro este
mismo tipo de células pueden dar paso al desarrollo de un embrión normal mediante el
proceso conocido como embriogénesis somática (ES) (Naumova, 1992; Yang y Zhang,
201 O; Zimmerman, 1993).
El proceso de ES atraviesa por una serie de cambios morfológicos y moleculares a través
de eventos de desdiferenciación celular, activación de la división celular, reprogramación
fisiológica y del metabolismo, así como cambios en los patrones de expresión genética.
Recientemente este proceso ha empezado a ser objeto de estudio para determinar la
participación de mecanismos epigenéticos en la formación de las estructuras
embriogénicas (Lardet et al., 2009; Yang y Zhang , 201 O; Yeung, 1995; Birchler et al.,
2011 ; Mohn y Schubeler, 2009). Si bien la ES es utilizada como una herramienta
biotecnológica para la propagación masiva, la conservación in vitro del germoplasma y el
mejoramiento genético, es sin duda también una valiosa herramienta para la investigación
científica (Griga, 2000; De Smet et al. , 2010; Zimmerman, 1993; Yang y Zhang, 2010).
29

CAPITULO 11
El proceso de diferenciación celular por la que atraviesa el embrión somático durante las
transiciones de un estadio a otro se ven fuertemente influenciado por mecanismos
epigenéticos (Eckardt, 2006; Zhang et al., 2013). Entre los mecanismos epigenéticos
estudiados durante la ES están la metilación del ADN y las modificaciones
postr.aduccionales en las histonas ryvu et al. , 2010; Deal y Henikoff, 2011; Nie-Gan et al.,
2013). Sin embargo, el tercer mecanismo epigenético ejercido por los pequeños ARNs en
este proceso no ha sido estudiado al nivel que permita entender su participación durante
la ES. Los miARN, una clase de ARNs pequeños, tienen un tamaño de 17-25 nt, han sido
identificados como moléculas esenciales para la diferenciación celular y el proceso de
desarrollo y germinación del embrión . Se han encontrado algunos miARNs, con
características conservadas evolutivamente en varias especies vegetales, involucrados en
el control de procesos biológicos tales como el ciclo del desarrollo vegetal, la floración, la
morfogénesis, la organogénesis y el desarrollo de órganos florales (Rhoades et al. , 2006;
Chen et al. , 2009).
El papel que ejercen los miARNs en el proceso de maduración y desarrollo de la
embriogénesis ha sido el centro de atención de varios grupos de investigación en los
últimos años (Rhoades et al, 2011 ; Zhang et al., 2006; Reinhart et al., 2002). Si bien se ha
logrado relacionar a algunos de los miARNs conservados con el proceso de
embriogénesis, obtener ciertos patrones sobre la distribución de estos miARNs e incluso
obtener nuevos candidatos que podrían estar relacionados con el proceso de
diferenciación celular y desarrollo embriogénico, aún no se ha podido elucidar el
mecanismo de su participación en dicho proceso. En esta parte de la investigación se
realizó la búsqueda in silico de los miARNs y el análisis de expresión de algunos de ellos
que se han reportado activos durante el proceso embriogénico en plantas.
2.2 MATERIALES Y MÉTODOS
2.2.1 Cultivo in vitro de Coffea canephora
Las plántulas de C. canephora fueron mantenidas y cultivadas in vitro en medio MS
(Murashige y Skoog, 1962), suplementado con 550 IJM de myo-inositol , 15 IJM de tiamina
HCI, 206.33 IJM de L-cisteina, 16.24 IJM de ácido nicotínico, 87.67 mM de sacarosa y
30

CAPITULO 11
0.25% (p/v) de gel Rite. El pH del medio fue ajustado a 5.8. Las plántulas fueron
mantenidas bajo condiciones de fotoperiodo (16h luz /8h oscuridad), a una temperatura de
25 ± 2 ·e y sub-cultivadas a medio fresco cada 35 días.
2.2.2 Inducción de la embriogénesis somática a partir de explantes foliares de Coffea canephora
Para el proceso de inducción de la ES, las plántulas fueron pre-acondicionadas por 14
días utilizando el medio de cultivo descrito previamente, modificando la concentración del
ácido nicotínico a 8.12 1JM, y agregando ANA, (0.541JM) y KIN (2.321JM) bajo las mismas
condiciones de cultivo. La inducción de la ES, fue llevada a cabo siguiendo el protocolo
propuesto por Quiroz-Figueroa (2006b). Brevemente, a partir de los primeros pares de
hojas, se obtuvieron discos foliares con un área de aproximadamente 0.25 cm2. Un total
de cinco explantes foliares fueron colocados en matraces Erlenmeyer con 50 mL de
medio Yasuda modificado, adicionado con benciladenina (BA, 5 IJM) (Hatanaka et al.,
1991 ). Los explantes fueron incubados en condiciones de oscuridad a 25 ± 2 oc y
mantenidos en agitación a 55 rpm por 56 días.
2.2.3 Extracción de los ARNs pequeños
Los ARNs pequeños fueron extraídos a partir de 100 mg de cada uno de los siguientes
tejidos vegetales, explantes foliares colectados a los O días de inducción, masa pro
embriogénica (Mp, colectada a los 21 días posteriores al proceso de inducción),
embriones somáticos de los estadios globular (Gio), corazón (Cor), torpedo (Tor) y
cotiledonar (Cot), aislados al dia 56 dpi. También se analizaron embriones cigóticos
colectados de plantas cultivadas en los jardines del Centro de Investigación Científica de
Yucatán.
La extracción se realizó utilizando el protocolo de cloruro de litio reportado previamente
por Rosas-Cardenas et al., (2011 ), con ciertas modificaciones para C. canephora. En
resumen, el tejido vegetal fue macerado en nitrógeno líquido con 2% de
polivinilpolipirrolidona (PVPP) (p/p) hasta la obtención de un polvo fino . El tejido
pulverizado fue homogenizado con 500 IJL del amortiguador de extracción [1 00 mM Tris
HCI (pH 9.0), 1% dodecil sulfato de sodio (SOS), 100 mM cloruro de litio (liCI) y 1 O mM de
ácido etilendiaminotetraacético (EDTA)] y 500 IJL de una solución saturada de fenol pH
31

CAPITULO 11
8.0, e incubado por 5 min a 60 oc. Posteriormente la mezcla fue centrifugada a 14,000
rpm por 1 O minutos a 4 oc y la fase superior acuosa fue transferida a un tubo nuevo.
Inmediatamente, se le adicionaron 600 IJL de cloroformo-alcohol isoamílico (24: 1, v/v) y la
mezcla se centrifugó a 11 ,500 rpm durante 1 O m in a 4 °C. Posteriormente, 350 IJL de la
fase superior se incubaron a 65 oc por 15 min. Finalizado el tiempo de incubación , se
adicionaron 60 IJL de NaCI 5 M y 73 IJL de polietilenglicol 8000 al 40%. La mezcla se
almacenó a -20 oc por 50 m in y después se centrifugó a 11,500 rpm durante 1 O m in a 4
°C. Seguidamente se tomaron 300 IJL de la fase superior y se le adicionaron 600 IJL de la
mezcla fenal-cloroformo-alcohol isoamílico (25:24:1, v/v/v) , la mezcla fue centrifugada a
11 ,500 rpm durante 1 O m in a 4 °C. Finalizada la centrifugación , se tomaron 300 IJL de la
fase superior y se le adicionaron 50 IJL de acetato de sodio 3 M (pH 5.2) y 1,700 IJL de
etanol absoluto frío. La mezcla se dejó precipitar por 15 horas a -20 oc y después fue
centrifugada a 11 ,500 rpm por 15 m in a 4 oc. El sobrenadante fue desechado y la pastilla
se dejó secar por 60 mina temperatura ambiente. Finalmente, la pastilla fue resuspendida
en 25 IJL de agua estéril libre de ARNasas y se realizó el análisis de la pureza y calidad
del ARN mediante la medición de absorbancia en los parámetros A260/A280 y
A260/A230.
2.2.4 Análisis electroforético de miARNs
El perfil electroforético de los pequeños ARNs de C. canephora se realizó utilizando geles
desnaturalizantes de urea/poliacrilamida al 12.5%. Los geles fueron preparados
mezclando 5 mL de la solución para geles al12.5% [acrilamida:bisacrilamida (19:1 Biorad)
y 8 M de urea], 25 IJL de tetrametiletilendiamina (TEMED) y 5 IJL de persulfato de amonio
(PSA) al 20%, dejando polimerizar los geles por 30 min a temperatura ambiente y
almacenados a 4 oc por 48 h. La preparación de las muestras partió de 3 IJg de ARN de
baja masa molecular añadiendo 0.3% (v/v) de amortiguador de carga (98% formamida, 10
mM EDTA, pH 8.0, 1 mg/mL xileno-cianol , 1 mg/mL de azul de bromo-fenal) más un
volumen de agua libre de ARNasas. Las muestras fueron incubadas a 65 oc durante 5
min para desnaturalizar el ARN y entonces fueron colocadas en hielo por 5 min. Los
pocillos de los geles fueron lavados introduciendo 3 mL de 0.5X de TBE (0.9 M Tris-HCI,
pH 8.0, 0.9 M ácido bórico y 2 mM EDTA) utilizando una jeringa para retirar el exceso de
PSA. La corrida electroforética se llevó a cabo por 2 h a 80 V utilizando como
amortiguador de corrida TBE 0.5X. Posteriormente, los geles fueron retirados de los
32

CAPITULO 11
cristales y teñidos con 1% de Syber-gold en 1 O mL de TBE 0.5X por 30 m in a temperatura
ambiente y realizando un lavado final con agua bidestilada por 5 min. La visualización de
las bandas fue realizada utilizando el equipo Gel Doc XR (Bio-Rad).
2.2.5 Análisis in silico de secuencias y diseño de sondas para los microARNs de Coffea canephora
Los miARNs que se consideraron analizar en este trabajo fueron a partir de una búsqueda
exhaustiva en artículos de investigación publicados hasta el momento en procesos como
la ES, EC y la respuesta a estrés in vitro. Posteriormente, las secuencias maduras de los
miARNs candidatos fueron buscadas en la base de datos miRBase
(http://www.mirbase.org), seleccionando las secuencias de los miARN maduros de las
e~pecies Vitis vinífera, Solanum lycopersicum, G/ycina· max, Popu/os trichocarpa, M.
truncatu/a, A. thalíana y O. sativa. Posteriormente, se realizó un alineamiento de
secuencias utilizando la plataforma de datos psRNATarget: A Plant Small RNA Target
Analysis Server (http://plantgrn.noble.org/psRNATarget/). A partir de este análisis, las
secuencias que presentaron mayor similitud fueron seleccionadas para el estudio en C .
. canephora. Las sondas seleccionadas. para nuestro estudio se enviaron a sintetizar y se
enlistan en el Cuadro 2.1.
Cuadro 2.1. Lista de sondas sintetizadas correspondientes a secuencias maduras de miARNs
m1ARNs Secuencta 5 '~3 ' moARNs Secuencia 5'-3 '
SnU6 5 "AGGGGCCATGCTAATCTTCTC3" miR397 S"TCATCAACGCTGCACTCAAT3'
5 '85 5'ACGTCTGCCTGGGTGTCACAA3' m iR5158 5' ATTCATCCTCATCCCAGTGGCTCA3"
miR1524 s· GGAGTTCCTTCCTCGGACTCG3" miR393 s· ATCAATGCGATCCCTTTGGAA 3'
miR157a S" GTGCTCTCTATCTTCTGTCAA3' miR400 5' TGACTTATAATACTCTCATA 3'
miR165a 5'GGGGGATGAAGCCTGGTCGGA3' miR778 5' TGTACATAAACCAAGCCA 3'
miR2119 5' TTCCCCTACAACTCCCTTTGA3' m iRB24 S'TTCTCACAAATGGTCTA 3'
miR3623a 5'TGGTGCTTGGATGAACTTGTGA3' miR399 S' GCAAATCTCCTTTGGCA 3'
miR3623b 5'ATGCAAATTCGTCCAAGCACCA3' m iRB27t 5' AATCTACTGGTAGTTGTTT 3'
miR3629b 5'TCCTACATTTTCTCAGCAGCC3' m iR827c 5' AATCTACTGGTAGTTGTTTG 3'
miR3635b 5'CAGGCATGTGTGGGACATAAT3' miRB19 5' AAAGTCTTATAACCTGA 3'
miR3636b 5'CTCCGACTTGCTTCTCCGACAGAC3' miR472 S'GGATGGGTGGAGTAGGGAAAA3'
m iR390 5'GGCGCTATCCCTCCTGAGCTT3' m iR2199 5' ACTATGTGAT CGTGCCTAGTC3'
m iR395 5'GGAGTTCCCCCAAACACTTCAG3' miR1862 S' TCCCAAAATAAACCAACCTCGT3'
m iR398 5'AAGGGGTGACCTGAGAACACA3' miR449 S'CCAGCTAACATACACTGCCA3'
miR482 5'TAGGAATGGGTGGAATTGGAAA3' m1R164 5T GCACGTGCCCTGCTTCTCCA3'
miR535 5'GCGTGCTCTCTCTCGTTGTCA3' miR154 5'CGAAGGGCAACACGGATAACCTA3'
miR822 5'CATGTGCAAATGCTTCCCGCA3' miR166 5'GGGGAATGAAGCCTGGTCCGA3'
m iR169 5'CCGGCAAGTCATCCTTGGCTG3' miR319 5'AGGAGCTCCCTTCAGTCCAAA3'
miR171 5'GATATTGGCACGGCTCAATCA3' miR156 5'GTGCTCTCTATCTTCTGTCAA3'
miR159 5'TAGAGCTCCCTTCAATCCAAA3' miR170 S'GATATTGACACGGCTCAATCA3'
m iR160 5'TGGCATACAGGGAGCCAGGCA3' miR172 S' ATGCAGCATCATCAAGATTCT3'
m iR168 5'TTCCCGACCTGCACCAA GCGA3' miR158 5'TGCTTTGTCTACATTTGGGA3'
m iR167 5'TAGATCATGCTGGCAGCTTCA3' miR430 S'TTACCCCAACATGTAGCACTTA3'
m iR163 5'ATCGAAGTTCCAAGTCCTCTTCAA3' miR427 S'GCCCAAAACAGAAAGCACTTT3'
m iR396 s'CAGTTCAAGAAAGCTGTGGAA3' miR309 S'CAAAACCGAACAAGGTTTGTCG3'
33

CAPITULO 11
2.3 RESULTADOS
2.3.1 Inducción, desarrollo morfológico y rendimiento de la ES en Coffea canephora
Para realizar el análisis de los miARNs durante el proceso de formación y desarrollo de
los embriones somáticos en C. canephora, como modelo de estudio, a partir de explantes
foliares sometidos al proceso inducción fue preciso hacer el seguimiento de la respuesta
al proceso embriogénico.
El proceso de la ES en C. canephora fue monitoreado durante 56 días (Figura 2.1 ).
Durante los primeros 14 días posteriores a la inducción, los explantes mostraron las
señales de cicatrización y engrosamiento en las heridas que son típicas de este proceso.
También se pudieron observar pequeñas estructuras dispersas, que indicaron el
comienzo de la diferenciación celular, como se muestra en la figura 2.1 D. Posteriormente,
en los días 21 y 28 dpi, se observó un aumento en la proliferación celular, como resultado
de una serie de divisiones celulares que caracterizan la diferenciación celular durante la
embriogénesis, al mismo tiempo que también se forma una masa Mp. Cuando han
transcurrido siete semanas de la inducción de la ES, se puede apreciar la formación de
embriones en el estadio globular. A partir de este momento se da una formación contínua
de los diferentes estadios embriogénicos, corazón, torpedo y cotiledonar. Durante los días
42 y 49 dpi se puede observar una mezcla de todos los estadios embriogénicos.
Finalmente, en el día 56 dpi, el explante se encuentra cubierto de estructuras
embriogénicas.
34

CAPITULO 11
Figura 2.1. Proceso de ES en Coffea canephora. (A) Preacondicionamiento de plántulas por catorce días en medio MS + ANA y KIN. (8) hojas primordiales. (C) explantes foliares para la inducción de ES y (D) Seguimiento del proceso embriogénico cada 7 días después de la inducción de la ES (dpi) hasta el día 56 dpi.
El análisis, aislamiento y cuantificación de los embriones somáticos en el explante
después de 56 dpi permitió determinar que se producen alrededor de 100 embriones
somáticos por explante, divididos de la siguiente manera, 20, 38, 2.5 y 15 para los estadios
Glo, Cor, Tor y Cot, respectivamente (Figura 2.28). Los embriones en estadio cotiledonar
fueron puestos a germinar, con el fin de determinar su viabilidad. Todos los embriones
germinaron y produjeron plantas sanas (Figura 2.2A).
35

CAPITULO 11
40
0 Estadios embriogénicos
Figura 2.2. Aislamiento y cuantificación de los embriones somáticos en C. anephora. (A) Los embriones somáticos aislados a los 56 dpi. Los embriones somáticos del estadio cotiledonar fueron colocados en medio MS para su conversión a plántulas. (B) Cuantificación de los embriones somáticos por explante 56 dpi. Las barras representan la cuantificación de tres réplicas biológicas. Las letras sobre las barras representan la diferencia significativa con una P>0.05 utilizando la prueba de Tukey.
2.3.2 Aislamiento de los ARN pequeños y miARNs en el proceso embriogénico de C. canephora
Para abordar molecularmente el presente estudio se aislaron los ARNs pequeños. Se
tomaron 100 mg de cada una de las muestras mencionadas en materiales y métodos para
la realización de la extracción de los ARNs pequeños. En la figura 2.3 se enlistan los·
tejidos analizados en este estudio, así como la cantidad de individuos por tejido para la
extracción de ARNs pequeños. Por ejemplo, si se parte de hoja solo se utilizan tres
explantes para obtener la cantidad requerida pero si se parte de embriones globulares, se
requieren de 690 embriones para realizar una extracción , por lo que para realizar una
extracción de ARNs pequeños del estadio globular se requieren de 35 explantes en
promedio.
Aunque existen protocolos estandarizados para la extracción de ARN total , para la
extracción de los pequeños ARN se requirió estandarizar una metodología para un
enriqueciendo de los ARNs con tamaños de 17-30 nt. El análisis de eficiencia del
36

CAPITULO 11
protocolo utilizado en las muestras de C. canephora se determinó mediante el patrón de
bandeo en las corridas electroforéticas de dos diferentes marcadores moleculares; uno de
330 a 10 pb (M1; Figura 2.3) , que sirve para distinguir a los ARN pequeños totales y el
otro de 17-25 nt (M2; Figura 2.3) que es útil para ubicar a los miAR N que se encuentren
en ese rango de tamaño. Se observó un patrón de bandeo de 17-25 nt en las muestras
correspondientes a Cor, Tor y Cot, las cuales tuvieron una mayor abundancia en las
bandas, en comparación a las muestras de hoja, Mp, Glo y EC (Figura 2.3). Esto sugiere
que los ARNs pequeños de esas muestras están más íntegros que las de hoja, Mp, Glo y
EC.
Je~lantes
25 explantes
~690 emb one:s
-4 4 embriones
.. 110 embriones
~68 embrlon s
- 70 embriones
Figura 2.3. Perfil de bandeo de las extracciones de los ARNs pequeños. (A) Tejido necesario para obtener 100 mg de tejido de hoja (hoja), masa pro-embriogénica (Mp) , embrión globular (Gio), corazón (Cor), torpedo (Tor), cotiledonar (Cot), embrión cigótico (Cig) . (B) Corrida electroforética en geles desnaturalizantes de poliacrilamida al12.5% de las muestras de hoja, Mp, Glo, Cor, Tor, Cot y Cig. M1 y M2 h son los marcadores moleculares de 330 a 10 pb y de 17 a 25 nt, respectivamente.
2.2.3 Búsqueda in silico de los miARNs presentes durante el proceso embriogénico de C. canephora
Para determinar los posibles miARNs candidatos que pudieran estar participando en la ES
de C. canephora se utilizaron las bases de datos miRBase (http://www.mirbase.org) y
Plant Small RNA Target Analysis Server (http://plantgrn.noble.org/psRNATarget/). Se
37

CAPITULO 11
obtuvieron 47 secuencias de miARNs que pudieran estar involucradas en el proceso de la
ES. En esta búsqueda se identificaron secuencias conservadas para C. canephora , de los
miR156, miR158, miR159, miR164, miR166, miR167, miR168, miR171 , miR390, miR397,
miR398. Asi también se identificaron otras secuencias en C. canephora, que han sido
estudiadas en funciones biológicas como la desdiferenciación y diferenciación celular, el
desarrollo vegetativo y el estrés in vítro, los cuales hasta el momento no habían sido
evaluados en el proceso de la ES (Cuadros 2.2; 2.3 y 2.4).
2.3 DISCUSIÓN
La ES es una valiosa herramienta para estudiar los procesos celulares, bioquímicos,
epigenéticos y moleculares que ocurren durante la _inducción , la transición entre estadios
de desarrollo y el desarrollo de embriones somáticos (Kiran y Thorpe, 1995b; De Smet et
al. , 2010; Yang y Zhang , 2010; Zimmerman , 1993).
Durante mucho tiempo el género Coffea ha sido objeto de diversos estudios para la
obtención de protocolos eficientes y reproducibles de multiplicación a gran escala. Los
estudios realizados para determinar el mejor tipo de explante (García y Menéndez, 1987) ,
las condiciones y medio de cultivo (Samson et al. , 2006) y los análisis durante el
desarrollo de estructuras embriogénicas (Nassuth et al. , 1980; Quiroz-Figueroa et al. ,
2002), han permitido optimizar la eficiencia y el desarrollo embriogénico de Coffea, así
como también , se ha logrado determinar algunos de los factores que intervienen en la
capacidad de diferenciación celular y el papel clave de ciertos reguladores del crecimiento
que adicionados en la etapa de pre-tratamiento mejoran y optimizan el proceso de
inducción , así como la ES (Carneiro , 1997; Ayii-Gutiérrez et al. , 2013; Hatanaka et al. ,
1991 ; Quiroz-Figueroa et al. , 2001 ). En este trabajo se comprobó la eficiencia
embriogénica que presentan los explantes de C. canephora (Figuras 2.1 y 2.2). También
se analizó el posible papel que podrían estar ejerciendo los ARNs pequeños, como
mecanismo epigenético, durante el proceso embriogénico.
Se sabe que la regulación transitoria ejercida por los ARNs pequeños es un paso clave en
los mecanismos epigenéticos que regulan procesos como la gametogénesis, la
fertilización y el desarrollo de embriones (Chuck et al., 2009) . Estas pequeñas moléculas
no solo han sido relacionadas con el mantenimiento de la integridad genómica cigótica, en
38

CAPITULO 11
estudios recientes se determinó que existe una correlación entre su funcionamiento y el
origen biogénico de los ARNs pequeños que determinan su modus operandi entre
diversos grupos taxonómicos; inclusive, se ha propuesto que, debido a determinados
modos de acción, podrían regular la herencia de las marcas en las histonas contribuyendo
directamente en la herencia epigenética en las plantas superiores (Schwach et al. , 2009;
Xie et al., 2004; Zhang et al. , 2013; Llave et al., 2002a) . En conjunto, la importancia sobre
el conocimiento de estos mecanismo epigenéticos, durante el proceso de la ES, radica en
que permite relacionar la regulación en la actividad transcripcional de ciertos genes,
debido a estados conformacionales específicos de la cromatina, que restringen o permiten
la expresión genética, con la metilación del ADN.
Por otro lado, las moléculas pequeñas de ARN, entre las que destacan los ARNs
pequeños de interferencia (siARN) y los microARNs (miARNs), son elementos
moduladores del mecanismo epigenético, pues la metilación de las histonas y la
metilación del ADN y en consecuencia la remodelación de la cromatina, podrían estar
regulados a través de estos ARNs pequeños (Zhang et al., 2013; Tariq y Paszkowski,
2004). Recientemente, utilizando herramientas de la genómica comparativa se ha
determinado la presencia de miARNs pequeños nuevos conservados en C. canephora.
Utilizando la base de datos para EST de Solanaceae Genome Netwoks se identificaron un
total de 42 genes blanco potenciales. La mayoría de esos genes codifican para factores
de transcripción que parecen participar en el crecimiento, desarrollo y en las respuestas al
estrés (Nellikunnuma y Chandrashekar, 2012). Utilizando un acercamiento similar Akter et
al. , (2014) identificaron varios miARNs potenciales en C. arabica y también determinaron
seis posibles genes blanco en A. thaliana para estos miARNs. Un caso aparte lo forman
los miARNs provenientes del ARN de los intrones. En C. canephora, recientemente se ha
descubierto uno de estos miARNs que regula la expresión de los genes que codifican
para la N-metiltransferasa, una enzima que participa en la biosíntesis de la cafeína. El
miARNs pequeño proviene de un intrón del gen que codifica para la teobromina sintasa
(Mohanan et al. , 2013).
Así mismo, en el presente trabajo se encontró la presencia de ARNs pequeños con
tamaños entre 17 a 25 nt de longitud durante todo el proceso de inducción y desarrollo de
la ES en C. canephora (Figura 2.3). Ha tomado más de 20 años de investigación para
39

CAPITULO 11
elucidar la función de los ARNs pequeños no codificantes en el proceso del desarrollo y
maduración de embriones cigóticos y somáticos en diferentes especies vegetales.
Estudios en A. thaliana (Schwarz et al. , 2008; Willmann et al. , 2011 ), M. truncatu/a (Eyles
et al. , 2013), V. orange (Xiao-Meng et al. , 2011), L. /eptolepis (Zhang et al. , 2012) y D.
logan (Lin y Lai , 2013) , han mostrado la función los miARNs involucrados en el desarrollo
de las estructuras embriogénicas, así como la importancia de los otros componentes,
tales como Dicer, Argonauta y HASTY, que participan en la biogénesis de las diferentes
clases de ARNs pequeños en las plantas, pues determinan en gran parte la funcionalidad
de estas moléculas (Wilbert y Yeo, 2011 ; Chen, 2005; Nonogaki, 201 O) .
El análisis del proceso de los miARNs ha ampliado el panorama sobre el papel en la
regulación que tienen estas moléculas en diferentes procesos biológicos (Cuadro 2.2, 2.3
y 2.4). Los microARNs: miR156, miR159, miR164, miR166, miR167, miR168, miR171 ,
miR390, miR397, miR398 y miR169 que val idamos experimentalmente en esta
investigación, se les ha relacionado con el proceso de la ES en C. orange, L. /eptolisis y
M. truncatula , lo que sugiere que se necesita cierta regulación por estos miARNs para que
se lleven a cabo los eventos biológicos implicados en la inducción y diferenciación celular
de la ES; asimismo, también se ha demostrado que al perderse la regulación
proporcionada por los miARNs, miR164/166/397, se puede inhibir parcial o totalmente el
desarrollo embriogénico (Xiao-Meng et al. , 2011; Zhang et al., 2012; Zhou et al., 2008;
Eyles et al. , 2013).
Por otra parte, con la utilización de diversas herramientas, provenientes de la genómica
comparativa, se ha avanzado en las predicciones de nuevos miARNs en diferentes
especies. Los estudios computacionales realizados en C. canephora y C. arabica han
identificado miARN conservados relacionados a 12 familias de miARNs en especies como
S. /ycopersicum, V. vinífera, A. tha/iana, G. max, P. trichocarpa y O. sativa (Nellikunnuma
y Chandrashekar, 2012; Akter et al. , 2014) . Por ello, para la síntesis de las sondas de los
miARNs analizados en esta investigación , se usaron las secuencias reportadas en estas
especies. Si bien , la mayoría de los miARNs se encuentran relacionados con factores de
transcripción que regulan el crecimiento y la respuesta a extremos ambientales, también
existen otros miARNs, como miR309, miR400, miR427, miR430 y miR2119, que se están
estudiando en este trabajo , debido a que se sabe que participan en el desarrollo
40

CAPITULO 11
embrionario en los mamíferos y en alg~nos vertebrados, y han sido analizados durante el
desarrollo de las plat:ltas, y deseamos determinar si estos miARNs podrían estar
involucrados en el desarrollo embrionario de las plantas (Sunkar y Zhu, 2004; Zhu et al.,
2008; Sunkar y Zhu, 2004).
Cuadro 2.2. miARNs conservados e involucrados en el desarrollo de las plantas y su papel
durante el proceso de ES.
miARNs Función Proteína codificada
Referencias por el gen diana
Transición de la fase juvenil a la Squamosa prometer- (Jiao et al. , 2010; Wu y fase adulta, desarrollo de raíz, miR156
maduración de EC ~ inducción de binding protein Poethig, 2006; Zhang
la ES. (SBP) et al., 2012)
Desarrollo de órganos florales,
miR159 germinación de semillas y MYB protein
(Achard et al. , 2004; formación de embriones en estadio Reyes y Chua, 2007) globular. Desarrollo de brotes, formación de NAC Domain (Kim et al. , 2009;
miR164 meristemos apicales y regulación containing proteins, Raman et al., 2008; en la transición de estadios CUC y GBOF-1- Sanan-Mishra et al., embriogénicos LIKE 2013) Formación de meristemos
miR166 apicales, desarrollo vascular, HD-ZIP transcription (Zhou et al., 2007; polaridad de hojas y el desarrollo factor Williams et al. , 2005) en la ES
Desarrollo de órganos Auxin response (Kinoshita et al. , 2012a; miR167 reproductivos, fertilización y factor (ARF's) Wu et al. , 2006)
desarrollo de los estadios en la EC
miR168 Modula la biogénesis de miARNs, Argonaute (AG01)
(Vaucheret et al. , 2006; el desarrollo de la ES Lin y Lai , 2013)
Desarrollo de flores, diferenciación GRAS domain or (Zhang et al. , 201 O; miR171 celular, regulación en el patrón del SCARECROW- like
sistema radicular proteins (SCL) Llave et al., 2002b)
Estrés in vitro, regula el ciclo de (Shen et al. , 2009;
miR390 desarrollo de raíces laterales y TAS3 and (ARFs) Marin et al. , 201 O; modula la formación del estadio
globular en la ES Yoon et al., 2010)
Regula la respuesta ante los
miR397 extremos por sequía, salinidad y Laceases and beta-6 (Sunkar y Zhu, 2004)
ácido abscísico (ABA) y la tubulin diferenciación celular Tolerancia a estrés oxidativo y ,POr Copper superoxide
miR398 deficiencia de nutrientes y modula dismutases (Sunkar et al. , 2006) el proceso de diferenciación celular (cds1,cds2, ccs1, y transición embriogénica cox5b-1)
41

CAPITULO 11
Cuadro 2.3. miARNs divergentes en el desarrollo de las plantas y su papel durante el proceso embriogénico.
Proteína miARNs Función codificada por el Referencias
gen blanco
Desarrollo de la morfogénesis en (Rhoades et al. , 2002;
miR157 hojas, semillas, regula los
Sbp-box Gandikota et al. , 2007; tiempos de floración y las fases Wang et al ., 2001 ; Kim et de transición vegetativa al. , 2012)
Regula la iniciación de raíces ARF10, ARF16;
(Mallory et al. , 2005; miR160 laterales, desarrollo de semilla y Nonogaki, 201 O; Qiao et
biosíntesis de las auxinas ARF17
al. , 2012)
Desarrollo y morfología de hoja, HD-Zip 111 gene, (Mallory et al. , 2005; Zhou
miR165 et al. , 2007; Williams et al. , el meristemo y las semillas phb, phv
2005) Desarrollo de la floración , regula
miR169 la respuesta ante estres por ABA ABRE,NF-YA (Un ver et al. , 201 O) y la proliferación celular Patrones de floración, regula la
APETALA-2, (Zhang et al. , 201 O; Zhu et miR172
diferenciación celular y el cambio RAP2.6,toe1, toe2, al. , 2009; Glazinska et al.,
de estadios vegetativos de las toe3,ids1, gloy15 2009)
plantas
Desarrollo de la semilla y (Ori et al. , 2007; Schommer et al. , 2008;
miR319 morfología de las hojas, regula la Tcp4, lanceolate,
Addo-Quaye et al. , 2009; respuesta ante los extremos de FamilyTFs
Li et al. , 2011 ; Ori et al. , salinidad y sequía 2007)
Desarrollo de hojas, raíces y (Chen et al. , 2011 ; Si-
TAAR, tir1 , afb2 y Ammour et al. , 2011 ; miR393 brotes apicales, modula la
afb3 Robert-Seilaniantz et al. , biosíntesis de auxinas 2011)
miR396 Regula el desarrollo y morfología Gfr, WRKY,
(Xia et al. , 2012b; de las hojas Giacomelli et al. , 2012) Regula la diferenciación celular y (Gar et al. , 2011 ; Li et al. ,
miR535 el estrés in vitro de cultivos NF-YA (HAP2) 2009)
micropropagados
42

CAPITULO 11
Cuadro 2.4. miARNs divergentes involucrados en el desarrollo de las plantas y extremos
ambientales.
Proteína miARNs Función codificada por el Referencias
gen diana
miR158 Regula el desarrollo de la ------ ( Rhoades , 2012; Reinhart et al. , planta 2002; Rhoades et al. , 2002)
miR309 Modula la programación (Bushati et al. , 2008) celular -------
Regula la respuesta ante miR395 los estreses de salinidad, ------- (Lu et al. , 2008)
sequía, y sul~atos
Regula el patrón de (Sunkar y Zhu, 2004; Zhu et al. , miR400 desarrollo y maduración de ------- 2008; Sunkar y Zhu, 2004)
embriones cigóticos
Regula la programación miR427 celular y desarrollo df1a, cxcr7, TDRD7 (Bushati et al. , 2008)
embriogénico
Regula la maduración del miR430 EC y el desarrollo del ------- (Eyles et al., 2013)
embrión cotiledonar
Regula la floración,
miR482 expansión de pétalos y Sp/2, MYB, NAC, (Li et al. , 201 O) respuesta a infecciones NAM virales
Interfiere sobre el estado
miR827 de compactación de la ------- (Lu et al. , 2008) cromatina, y biosíntesis del (GA)
miR1524 Modula el de·sarrollo del
GAMYB (Subramanian et al. , 2008) sistema radicular
Regula la maduración del (Eyles et al. , 2013; Naqvi et al., miR2199 EC y el desarrollo del ------- 2010)
embrión cotiledonar
43

CAPITULO 11
2.5 REFERENCIAS
Achard , P. , Herr, A. , Baulcombe, D.G., y Harberd, N.P. (2004). Modulation of floral development by a gibberellin-regulated microRNA. Development 131 , 3357-3365.
Addo-Quaye, C. , Snyder, J.A. , Park, Y.B ., Li , Y.F. , Sunkar, R., y Axtell , M.J. (2009). Sliced microRNA targets and precise loop-first processing of MIR319 hairpins revealed by analysis of the Physcomitrella patens degradome. RNA 15, 2112-2121 .
Akter, A. , Islam, M. , Mondal, S.l. , Mahmud, Z., Jewel, N.A. , Ferdous, S. , Amín, M., y Rahman , M. (2014). Computational identification of miRNA and targets from expressed sequence tags of coffee (Coffea arabica). Saudi Journal of Biological Sciences 21 , 3-12.
Ayii-Gutiérrez, B., Galaz-Avalos, R.M ., Peña-Cabrera, E., y Loyola-Vargas, V.M. (2013). Dynamics of the concentration of IAA and some of its conjugates during the induction of somatic embryogenesis in Coffea canephora. Plant Signaling and Behavior 8, e26998.
Birchler, J.A. , Gao, Z., Sharma, A. , Presting, G.G. , y Han, F. (2011) . Epigenetic aspects of centro mere function in plants. Current Opinion in Plant Biology 14, 217-222.
Bushati , N. , Stark, A. , Brennecke, J., y Cohen , S.M. (2008). Temporal reciprocity of miRNAs and their targets during the maternal-to-Zygotic transition in Drosophi/a . Current Biology 18, 501-506.
Cangahuala-lnocente, G.C., Villarino, A. , Seixas, D., Dumas-Gaudot, E., Terenzi , H., y Guerra, M.P. Differential proteomic analysis of developmental stages of Acca sel/owiana somatic embryos. Acta Physiology Plant 31 , 514. 2009.
Carneiro, M.F. (1997). Coffee biotechnology and its application in genetic transformation . Euphytica 96, 167-172.
Chen, R. , Hu, Z. , y Zhang , H. (2009). ldentification of microRNAs in wild soybean (Giycine soja). Journal of lntegrative Plant Biology 51 , 1071-1079.
Chen , X. (2005) . microRNA biogenesis and function in plants. FEBS Letters 579, 5923-5931 .
Chen , Z.H., Bao, M.L., Sun , Y.Z., Yang , Y.J., Xu , X.H., Wang, J.H., Han, N., Bian , H.W., y Zhu , M.Y. (2011 ). Regulation of auxin response by miR393-targeted transport inhibitor response protein-1 is involved in normal development in Arabidopsis. Plant Molecular Biology 77, 619-629.
Chuck, G. , Candela, H. , y Hake, S. (2009) . Big impacts by small RNAs in plant development. Current Opinion in Plant Biology 12, 81-86.
De Smet, 1. , Lau, S., Mayer, U., y Jurgens, G. (201 O) . Embryogenesis - the humble beginnings of plant life. The Plant Journal 61 , 959-970.
44

CAPITULO 11
Deal, R. B. y Henikoff, S. (2011 ). Histone variants and modifications in plant gene regulation . Current Opinion in Plant Biology 14, 116-122.
Eckardt, N.A. (2006). Genetic and epigenetic regulation of embryogenesis. The Plant Cell 18, 781-784.
Eyles, R.P., Williams, P.H. , Ohms, S.J. , Weiller, G.F., Ogilvie, H.A. , Djordjevic, M.A. , y lmin, N. (2013). microRNA profiling of root tissues and root forming explant cultures in Medicago truncatula . Planta 238, 91-105.
Gandikota, M. , Birkenbihl, R.P., Hohmann, S., Cardon , G.H., Saedler, H. , y Huijser, P. (2007). The miRNA 156/157 recognition element in the 3' UTR of the Arabidopsis SBP box gene SPL3 prevents early flowering by translational inhibition in seedlings. The Plant Journal 49, 683-693.
Gar, 0. , Sargent, D.J., Tsai , C.J., Pleban, T., Shalev, G., Byrne, D.H. , y Zamir, D. (2011). An autotetraploid linkage map of rose (Rosa hybrida) validated using the strawberry (Fragaria vesca) genome sequence. Plos One 6, e20463.
García, E. , y Menéndez, A. (1987). Embriogénesis a partir de explantes foliares del cafeto catimor. Café Cacao Thé 31 , 15-22.
Giacomelli , J.l., Weigel, D., Chan, R.L. , y Manavella, P.A. (2012). Role of recently evolved miRNA regulation of sunflower HaWRKY6 in response to temperature damage. New Phytologist 195, 766-773.
Glazinska, P. , Zienkiewicz, A., Wojciechowski , W., y Kopcewicz, J. (2009). The putative miR172 target gene In APETALA2-Iike is involved in the photoperiodic flower induction of lpomoea ni/. Journal of Plant Physiology 166, 1801-1813.
Griga, M. (2000). Morphological alterations in sterile mutant of Pisum sativum obtained vía somatic embryogenesis. Biología Plantarum 43, 161-165.
Hatanaka, T., Arakawa, 0 ., Yasuda, T. , Uchida, N. , y Yamaguchi, T. (1991). Effect of plant growth regulators on somatic embryogenesis in leaf cultures of Coffea canephora. Plant Cell Reports 10, 179-182.
Jiao, Y. , Wang , Y. , Xue, D. , Wang , J., Yan, M., Liu, G., Dong , G., Zeng, D., Lu, Z. , y Zhu, X. (2010). Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Natural Genetics 42, 541-544.
Kapinas, K., Grandy, R. , Ghule, P., Medina, R. , Becker, K., Pardee, A. , Zaidi , S.K., Lían, J., Stein , J. , Van Wijnen , A. , y Stein , G. (2013). The abbreviated pluripotent cell cycle. Journal of Cellular Physiology 228, 9-20.
Kim, J.H., Woo, H.R. , Kim, J., Lim, P.O., Lee, I.C., Choi , S.H., Hwang, D., y Nam, H.G. (2009). Trifurcate feed-forward regulation of age-dependent cell death involving miR164 in Arabidopsis. Science 323, 1053-1057.
45

CAPITULO 11
Kim, J.J. , Lee, J.H. , Kim , W., Jung, H.S., Huijser, P., y Ahn, J.H. (2012). The microRNA156- SQUAMOSA PROMOTER BINDING PROTEIN-LIKE3 module regulates ambient temperature-responsive flowering vía FLOWERING LOCUS T in Arabidopsis. Plant Physiology 159, 461-478.
Kinoshita , N., Wang , H., Kasahara, H. , Liu , J. , Mac-Pherson, C., Machida, Y. , Kamiya, Y., Hannah, M.A., y Chua, N.H. (2012). /AA-Ala resistant3, an evolutionarily conserved target of miR167, mediates Arabidopsis root architecture changes during high osmotic stress. The Plant Cell 24, 3590-3602.
Kiran, K.S. y Thorpe, T.A. (1995). Asexual embryogenesis in vascular plants in nature. Mexico: Current Plant Science and Biotechnology in Agriculture.
Lardet, L., Dessailly, F., Carron , M.P., Montero, P., y Monteuuis, O. (2009). lnfluences of aging and cloning methods on the capacity for somatic embryogenesis of a mature Hevea brasiliensis genotype. Tree Physiology 29, 291-298.
Li , H., Deng, Y., Wu , T. , Subramanian, S., y Yu, O. (2010). Misexpression of miR482, miR1512, and miR1515 increases soybean nodulation. Plant Physiology 153, 1759-1770.
Li , H., Zhang , Z., Huang, F., Chang, L. , y Ma, Y. (2009) . MicroRNA expression profiles in conventional and micropropagated strawberry (Fragaria x ananassa Duch.) plants. Plant Cell Reports 28, 891-902.
Li , Y. , Li , Ch. , Ding , G., y Jin , Y. (2011). Evolution of M/R159/319 microRNA genes and their post-transcriptional regulatory link to siRNA pathways. BMC Evolutionary Biology 11, 122.
Lin , Y., y Lai , Z. (2013). Comparative analysis reveals dynamic changes in miRNAs and their targets and expression during somatic embryogenesis in Longan (Dimocarpus longan Lour). Plos One 8, e60337.
Llave, C. , Kasschau, K.O. , Rector, M.A. , y Carrington, J.C. (2002a). Endogenous and silencing-associated small RNAs in plants. The Plant Cell 14, 1605-1619.
Llave, C., Xie, Z., Kasschau, K.O., y Carrington , J.C. (2002b). Cleavage of Scarecrow-like mRNA targets directed by a class of Arabidopsis miRNA. Science 297, 2053-2056.
Lu , S. , Sun, Y.H., y Chiang , V.L. (2008). Stress-responsive microRNAs in Populus. The Plant Journal 55, 131-151 .
Lu , Y., Gan, Q., Chi , X., y Qin, S. (2008). ldentification and characterization of microRNAs and their targets in grapevine ( Vitis vinífera). Agricultura! Sciences in China 7, 929-943.
Mallory, A.C., Bartel , O.P., y Bartel, B. (2005). MicroRNA-directed regulation of Arabidopsis AUXIN RESPONSE FACTOR17 is essential for proper development and modulates expression of early auxin response genes. The Plant Cell 17, 1360-
46

CAPITULO 11
1375.
Marin , E. , Jouannet, V. , Herz, A. , Lokerse, AS., Weijers, D., Vaucheret, H. , Nussaume, L., Crespi, M.O., y Maizel , A. (2010). miR390, Arabidopsis TAS3 tasiRNAs, and their AUXIN RESPONSE FACTOR targets define an autoregulatory network quantitatively regulating lateral root growth. The Plant Cell 22, 1104-1117.
Mohanan , S. , Gowda, K. , Kandukuri , S.V. , y Chandrashekar, A. (2013). lnvolvement of a novel intronic microRNA in cross regulation of N-methyltransferase genes involved in caffeine biosynthesis in Coffea canephora. Gene 519, 107-112.
Mohn, F. y Schubeler, D. (2009). Genetics and epigenetics: stability and plasticity during cellular differentiation. Trends Genetics 25, 129-136.
Murashige, T. y Skoog , F. (1962) . A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia Plantarum 15, 473-497.
Naqvi , A.R. , Haq, Q.M., y Mukherjee, S.K. (201 0) . MicroRNA profiling of tomato /eaf curl new delhi virus (tolcndv) infected tomate leaves indicates that deregulation of mir159/319 and mir172 might be linked with leaf curl disease. Virology Journal 7, 281-296.
Nassuth, A., Wormer, T.M., Bouman , F., y Staritsky, G. (1980) . The histogenesis of callus in Coffea canephora stem explants and the discovery of early embryoid initiation. Acta Botanica Neerlandica 29, 49-54.
Naumova, T. (1992). Apomixis in Angiosperms: Nucellar and lntegumentary Embryony. 1er Edition
Nellikunnuma, S.M. y Chandrashekar, A. (2012). Computational identification of conserved miRNA and their targets in Coffea canephora by EST Analysis. Dynamic Biochemistry. Process Biotechnology and Molecular Biology 6, 70-76.
Nonogaki , H. (201 0). MicroRNA gene regulation cascades during early stages of plant development. Plant and Cell Physiology 51, 1840-1846.
Ori , N., Cohen, A.R. , Etzioni , A. , Brand, A. , Yanai, O., Shleizer, S., Menda, N., Amsellem, Z., Efroni, 1. , Pekker, 1., Alvarez, J.P. , Blum, E., Zamir, D. , y Eshed , Y. (2007). Regulation of LANCEOLA TE by miR319 is requi red for compound-leaf development in tomate. Natural Genetics 39, 787-791.
Qiao, M. , Zhao, Z ., Song , Y. , Liu, Z. , Cao, L. , Yu , Y. , Li , S., y Xiang , F. (2012) . Proper regeneration from in vitro cultured Arabidopsis thaliana requires the microRNAdirected action of an auxin response factor. The Plant Journal 71 , 14-22.
Quiroz-Figueroa, F.R. , Fuentes-Cerda, C. , Rojas-Herrera, R., y Loyola-Vargas, V.M. (2002). Histological studies on the developmental stages and differentiation of two different somatic embryogenesis systems of Coffea arabica. Plant Cell Reports 20, 1141-1149.
47

CAPITULO 11
Quiroz-Figueroa, F.R., Méndez-Zeel, M., Larqué-Saavedra, A. , y Loyola-Vargas, V.M. (2001). Picomolar concentrations of salicylates induce cellular growth and enhance somatic embryogenesis in Coffea arabica tissue culture. Plant Cell Reports 20, 679-684.
Quiroz-Figueroa, F.R., Monforte-González, M., Galaz-Avalos, R.M., y Loyola-Vargas,V.M. (2006a). Direct somatic embryogenesis in Coffea canephora. In Plant cell culture protocols, Loyola-Vargas, V.M., y Vázquez-Fiota, F.A., eds. Totowa, New Jersey: Humana 111-117.
Quiroz-Figueroa, F.R. , Rojas-Herrera, R., Galaz-Avalos, R.M., y Loyola-Vargas, V.M. (2006b). Embryo production through somatic embryogenesis can be used to study cell differentiation in plants. Plant Cell Tissue and Organ Culture 86, 285-301.
Raman, S., Greb, T. , Peaucelle, A. , Blein , T. , Laufs, P., y Theres, K. (2008). lnterplay of miR164, CUP-SHAPED COTYLEDON genes and LATERAL SUPPRESSOR controls axillary meristem formation in Arabidopsis thaliana. The Plant Journal 55, 65-76.
Reyes, J.L. y Chua, N. (2007) . ABA induction of miR159 controls transcript levels of two MYB factors during Arabidopsis seed germination. The Plant Journal 49, 592-606.
Reinhart, B.J. , Weinstein, E.G. , Rhoades, M.W., Bartel , B., y Bartel, O.P. (2002). MicroRNAs in plants. Genes and Development 16, 1616-1626.
Rhoades, M.W., Bartel , O.P. , y Bartel, B. (2006). MicroRNAS and their regulatory roles in plants. Annual Review of Plant Biology 57, 19-53.
Rhoades, M.W. (2012). Conservation and divergence in plant microRNAs. Plant Molecular Biology 80, 3-16.
Rhoades, M.W., y Rhoades, J. (2011). Conservation and divergence in plant microRNAs. Plant Molecular Biology 80, 3-16
Rhoades, M.W., Reinhart, B.J ., Lim, L.P., Surge, C.B. , Bartel , B. , y Bartel, O.P. (2002). Prediction of plant microRNA targets. Cell 11 O, 513-520.
Robert-Seilaniantz, A. , MacLean, D., Jikumaru, Y. , Hill, L., Yamaguchi , S., Kamiya, Y. , y Jones, J.D. (2011). The microRNA miR393 re-directs secondary metabolite biosynthesis away from camalexin and towards glucosinolates. The Plant Journal 67, 218-231.
Rosas-Cardenas, F.F., Duran-Figueroa, N., Vielle-Calzada, J.P., Cruz-Hernandez, A. , Marsch-Martinez, N., y De Folter, S. (2011) . A simple and efficient method for isolating small RNAs from different plant species. Plant Methods 7, 4-11
Samson, N.P. , Campa, C. , Gal, L.L., Noirot, M. , Thomas, G., Lokeswari , T.S. , y Kochko, A. (2006). Effect of primary culture medium composition on high frequency somatic embryogenesis in different Coffea species. Plant Cell Tissue and Organ Culture 86,
48

CAPITULO 11
37-45.
Sanan-Mishra, N. , Varanasi , S., y Mukherjee, S. (2013). Micro-regulators of auxin action. Plant Cell Reports 32, 733-740.
Schommer, C. , Palatnik, J.F., Aggarwal , P., Chetelat, A. , Cubas, P. , Farmer, E.E., Nath, U. , y Weigel, D. (2008). Control of jasmonate biosynthesis and senescence by miR319 targets. Plos Biology 6, e230.
Schwach, F., Moxon, S., Moulton, V., y Dalmay, T. (2009). Deciphering the diversity of small RNAs in plants: The long and short of it. Briefing Functional Genomics Proteomics 8, 472-481 .
Schwarz, S., Grande, AV., Bujdoso, N., Saedler, H. , y Huijser, P. (2008). The microRNA regulated SBP-box genes SPL9 and SPL 15 control shoot maturation in Arabidopsis. Plant Mololecular Biology 67, 183-195.
Shen, D. , Wang , S., Chen, H., Zhu, Q.H ., Helliwell, C., y Fan, L.J. (2009). Molecular phylogeny of miR390-guided trans-acting siRNA genes (TAS3) in the grass family. Plant Systematics and Evolution 283, 125-132.
Si-Ammour, A. , Windels, D. , Bouldoires, E., Kutter, C. , Ailhas, J. , Meins, F., y Vazquez, F. (2011 ). miR393 and secondary siR NAs regulate expression of the TIR1/AFB2 auxin receptor clade and auxin-related development of Arabidopsis Leaves. Plant Physiology 157, 683-691 .
Subramanian, S. , Fu, Y., Sunkar, R. , Barbazuk, W.B. , Zhu, J.K., y Yu , O. (2008). Novel and nodulation-regulated microRNAs in soybean roots. BMC Genomics 9, 160.
Sunkar, R. , y Zhu, J.K. (2004). Novel and stress-regulated microRNAs and other small RNAs from Arabidopsis. The Plant Cell16, 2001-2019.
Sunkar, R. , Kapoor, A. , y Zhu, J.K. (2006) . Posttranscriptional induction of two Cu/Zn superoxide dismutase genes in Arabidopsis is mediated by downregulation of miR398 and important for oxidative stress tolerance. The Plant Cell 18, 2051-2065.
Tariq , M. y Paszkowski, J. (2004). DNA and histone methylation in plants. Trends in Genetics 20, 244-251.
Unver, T., Parmaks, 1., y Dündar, E. (201 0). ldentification of conserved micro-RNAs and their target transcripts in opium poppy (Papaver somniferum L.). Plant Cell Reports 29, 757-769.
Vaucheret, H., Mallory, A.C., y Bartel , O.P. (2006). AG01 homeostasis entails coexpression of MIR168 and AG01 and preferential stabilization of miR168 by AG01 . Molecular Cell 22, 129-136.
Wang, W.X., Gaffney, B., Hunt, A.G., y Tang, G. (2001). MicroRNAs (miRNAs) and plant development. In eLS, John Wiley and Sons, Ud.
49

CAPITULO 11
Wilbert, M.L. y Yeo, G.W. (2011). Genome-wide approaches in the study of microRNA biology. WIREs Systems Biology and Medicine 3, 491-512.
Williams, L. , Grigg , S.P., Xie, M., Christensen, S., y Fletcher, J.C. (2005). Regulation of Arabidopsis shoot apical meristem and lateral organ formation by microRNA miR166g and its AtHD-ZIP target genes. Development 132, 3657-3668.
Wu, G. , y Poethig, R.S. (2006). Temporal regulation of shoot development in Arabidopsis thaliana by miR156 and its target SPL3. Development 133, 3539-3547.
Wu, L. , Zhou, H., Zhang, Q., Zhang , J., Ni, F., Liu, C. , y Qi , Y. (2010). DNA methylation mediated by a microRNA pathway. Molecular Cell 38, 465-475.
Wu, M.F., Tian, Q., y Reed, J.W. (2006). Arabidopsis microRNA167 controls patterns of ARF6 and ARF8 expression, and regulates both female and male reproduction. Development 133, 4211-4218.
Xia, K., Wang , R. , Ou , X., Fang , Z. , Tian, C., Duan, J., Wang , Y., y Zhang, M. (2012) . OsTIR1 and OsAFB2 downregulation via OsmiR393 overexpression leads to more tillers, early flowering and less tolerance to salt and drought in rice. Plos One 7, e30039.
Xiao-Meng, Wu., Mei-Ya, L., Xiao-Xia, Ge., Qiang , Xu ., and Wen-Wu, G. (2011). Stage and tissue-specific modulation of ten conserved miRNAs and their targets during somatic embryogenesis of Valencia sweet orange. Planta 233, 495-505.
Xie , Z. , Johansen, L.K. , Gustafson, A.M ., Kasschau, K.O., Lellis, A.D. , Zilberman, D., Jacobsen, S.E., y Carrington, J.C. (2004). Genetic and functional diversification of small RNA pathways in plants. Plos Biology 2, E1 04.
Yang, X. y Zhang, XL. (201 0). Regulation of somatic embryogenesis inhigher plants. Critica! Reviews in Plant Sciences 29, 36-57.
Yasuda, T., Tahara, M., Hatanaka, T., Nishibata, T., y Yamaguchi , T. (1995). Clonal propagation through somatic embryogenesis of Coffea species. XVI Colloq. Sci. lnt. Coffee. Kyoto 392.
Yeung, E.C. (1995). Structural and development patters in somatic embryogenesis. In In Vitro Embryogenesis in Plants. Thorpe T.A. Ed : Kluwer Academic Publishers, Netherlans 205-24 7.
Yoon, E.K. , Yang , J.H., Lim, J., Kim, S.H., Kim, S.K. , y Lee, W.S. (2010). Auxin regulation of the microRNA390-dependent transacting small interfering RNA pathway in Arabidopsis lateral root development. Nucleic Acids Research 38, 1382-1391.
Zhang, B., Pan, X., Cannon , C.H., Cobb, G.P. , y Anderson, T.A. (2006). Conservation and divergence of plant microRNA genes. The Plant Journal 46, 243-259.
Zhang, H. , Chaudhury, A. , y Wu, X. (2013). lmprinting in plants and its underlying
50

CAPITULO 11
mechanisms. Journal of Genetics and Genomics 40, 239-247.
Zhang , J. , Zhang , S. , Han, S., Wu, T., Li , X. , Li, W. , y Qi , L. (2012). Genome-wide identification of microRNAs in larch and stage-specific modulation of 11 conserved microRNAs and their targets during somatic embryogenesis. Planta 236, 647-657.
Zhang, S., Zhou, J. , Han, S. , Yang, W., Li , W. , Wei , H. , Li , X ., y Qi , L. (2010). Four abiotic stress-induced miRNA famil ies differentially regulated in the embryogenic and nonembryogenic callus tissues of Larix Jeptolepis. Biochemical and Biophysical Research Communications 398, 355-360.
Zhou , G.K., Kubo, M., Zhong , R. , Demura, T. , y Ye, Z.H. (2007) . Overexpression of miR165 affects apical meristem formation , organ polarity establishment and vascular development in Arabidopsis. Plant and Cell Physiology 48, 391-404.
Zhou, Z.S. , Huang, S.Q., y Yang , Z.M. (2008). Bioinformatic identification and expression analysis of new microRNAs from Medicago truncatula . Biochemical and Biophysical Research Communication 374, 538-542.
Zhu, Q.H., Spriggs, A. , Matthew, L. , Fan , L., Kennedy, G. , Gubler, F. , y Helliwell , C. (2008) . A diverse set of microRNAs and microRNA-Iike small RNAs in developing rice grains. Genome Research 18, 1456-1465.
Zhu, Q.H. , Upadhyaya, N.M. , Gubler, F., y Helliwell , C.A. (2009). Over-expression of m iR 172 causes loss of spikelet determinacy and floral organ abnormalities in rice (Oryza sativa) . BMC Plant Biology 9, 149.
Zimmerman, J.L. (1993). Somatic embryogenesis: A model for early development in higher plants. The Plant Cell 5, 1411-1423.
51

\

CAPITULO 111
CAPÍTULO 111 ANÁLISIS DE LA EXPRESIÓN DIFERENCIAL DE miARNs
DURANTE LA EMBRIOGÉNESIS SOMÁTICA DE C. Canephora
3.1 Introducción
Los miARNs son moléculas pequeñas, no codificantes, que realizan un papel muy
importante en la regulación del desarrollo de las plantas. Con la creciente innovación
tecnológica como la secuenciación masiva y el desarrollo de herramientas
bióinformáticas, ahora se puede predecir la presencia de miARNs nuevos; utilizando este
acercamiento se ha dado un gran avance en el número de miARNs y su presencia en
varias especies de plantas (Thakur et al., 2011; Adai et al. , 2005; Wang et al., 2004;
Rhoades et al., 2002). Si bien biológicamente se ha llegado a comprender la biogénesis y
el mecanismos de acción de estas moléculas, el reto biológico que aun persiste gira
alrededor de la identificación y validación de las funciones que desempeñan estas
moléculas (Tretter et al., 2008; Rogers y Chen, 2013; Chen, 2005; Allen et al. , 2005) .
Entre los procesos que sabemos que regulan los miARNs se encuentran el desarrollo de
los órganos florales, la morfogénesis de hojas, la respuesta generada ante diversos
extremos ambientales y el desarrollo de los embriones (Mallory y Vaucheret, 2006; Pei et
al., 2013; Wang et al., 2001; Palatnik et al., 2003; Khraiwesh et al., 2012; Mallanna y
Rizzino, 201 0). Se ha evaluado ampliamente la expresión de estas moléculas y se ha
hecho evidente que existe una distribución espacio-temporal de sus patrones de
expresión lo que a su vez controla la activación y la represión del programa de desarrollo
de las plantas, creando así una base sustentable para una serie de predicciones sobre la
función de los miARNs y sus genes blancos (Llave et al. , 2002; Wang et al., 2005; Bonnet
et al. , 2004; Cuperus et al., 2011).
Durante los procesos de embriogénesis cigótica y somática ocurren eventos biológicos
que requieren de una adecuada programación en la expresión génica (Yang y Zhang ,
2010; Xu y Huang, 2014; Aquea y Arce-Johnson, 2008; Karami et al. , 2009). La
53

CAPITULO 111
importancia de la función de los miARNs durante el desarrollo y la maduración de los
embriones se describió inicialmente en embriones cigóticos de A. thaliana, en particular
de la enzima responsable de la biogénesis de los miARNs, Dicer-like1 (DCL-1). Se
descubrió que la sincronización del programa de maduración de los embriones se lleva a
cabo a través de la regulación por miARNs, controlando la represión de los reguladores
LEAFY COTYLEDON-2 (LEC2) y FUSCA-3, así como de los factores transcripcionales
ARABIDOPSIS 68-INTERACTING PROTEIN1-LIKE1 (ASIL 1) 1 ASIL2 y a la deacetilasa
de la histona HDA6/SIL 1 los cuales participan en los estadios tempranos de la
embriogénesis en Arabidopsis (Willmann et al., 2011 ).
En los últimos tres años, diferentes reportes sobre el papel de los miARNs durante el
proceso de EC en A. thaliana (Willmann et al. , 2011 ), P. teada (Oh et al., 2008) y L.
/eptolepis (Zhang et al., 2013) y durante la ES en M. truncatula (Eyles et al., 2013), L.
/eptolepis (Zhang et al., 2012), D. Logan (Lin y Lai, 2013), O. Sativa (Zhou et al., 201 O) y
V. orange (Xiao-Meng et al., 2011), muestran que existen un claro patrón de expresión de
los miARNs durante las etapas que comprenden el desarrollo embriogénico (Jenik et al.,
2007). Por ejemplo, se ha descubierto que durante la primera etapa de la ES en C.
sinensis, miR156 actúa sobre ocho genes blancos putativos, todos ellos codifican para
Squamosa promoter-binding proteins (SBP). Siete de esos genes son Squamosa
Promoter-Binding-like (SPL) e incluyen a los genes SPL2 (uno), SPL4 (uno), SPL 13 (uno),
SPL5 (dos) y SPL9 (dos) (Xiao-Meng et al., 2011). Hoy en día se sabe que una correcta
programación durante la etapa temprana del estadio de corazón resulta crucial para el
modelado del eje embrionario y del meristemo (Jenik et al. , 2007) . No obstante, el papel
que desempeñan los miARNs en la segunda etapa de la ES, es decir, durante la
transición de los estadios embriogénicos tempranos hasta la conversión completa al
estadio cotiledonar, no está clara.
En los últimos años, varios autores han estudiado el papel de miR156, miR159, miR164,
miR166, miR167, miR168, miR171, miR390, miR397 y miR398, clasificados como
miARNs conservados evolutivamente en diferentes especies, y su relación en la
modulación de genes blanco a través de su expresión diferencial en varios procesos del
desarrollo vegetativo (Rhoades, 2012). Algunos de estos miARNs regulan factores como
SEPALA TA (SPL), Argonauta (AG01) y Scarecrow-like (SCL) (Zhang et al., 201 0). Por
54

CAPITULO 111
otro lado miR159, miR162, miR164, miR390 y miR397 participan modulando la expresión
de NAC, MYB-protein, TAS3 y LACCASES (Zhang et al., 2012; Xiao-Meng et al., 2011).
Los miARNs miR160, miR166, miR167 y miR398 también participan en la regulación de
los estadios tardíos de la ES y actúan sobre los factores Auxin Response Factor (ARF's),
PHABULOSA (PHB) y LACCASES (Oh et al., 2008; Zhang et al., 2012). Finalmente, con
base en el estudio bioinformático realizado en C. canephora, en el que se determinaron
18 miARNs pertenecientes a 12 familias y 42 blancos potenciales (Nellikunnuma y
Chandrashekar, 2012) y utilizando el proceso embriogénico establecido en C. canephora
en esta investigación se estudiaron 47 miARNs como posibles candidatos de la regulación
del proceso embriogénico de esta especie.
3.2 Materiales y métodos
3.2.1 Recuperación y purificación de fragmentos pequeños (17 -25 nt) de ARN
La purificación de los fragmentos de ARN pequeños se realizó a partir de una corrida
electroforética con 25 ¡Jg del ARN de cada una de las muestras (Hj, Mp, Glo, Cor, Tor, Cot
y Cig). Posteriormente, las bandas de 17 - 25 nt fueron cortadas y colocadas en tubos
Eppendorf, triturando el gel con una varilla de vidrio en 200 !JL de agua bidestilada, hasta
obtener una mezcla homogénea. El volumen total de la mezcla fue incubada a 70 OC por
10 min, seguida de una centrifugación a 3,000 rpm por 2 min a 4 °C. Se tomó la fase
acuosa y el volumen final se llevó a 500 IJL con agua libre de ARNsas, se le adicionaron 8
!JL de glucógeno (5 mg/ml), 35 !JL de NaOAc 3 M (pH 5.2) y 900 !JL de etanol absoluto
frío. La mezcla se incubó por 1 ha -80 OC y se centrifugó a 7,000 rpm por 10 mina 4 °C.
El sobrenadante se descartó y la pastilla se dejó secar por 30 m in y se resuspendió en 1 O
¡JL de agua libre de ARNsas y almacenada a -20 OC hasta su uso.
3.2.2 Desfosforilación a partir de los fragmentos purificados de C. canephora
La reacción de desfosforilación de las muestras (Hj, Mp, Glo, Cor, Tor, Cot y Cig) se
realizó en los ARNs purificados, preparando reacciones con un volumen total de 30 IJL.
Cada mezcla se realizó a partir de 900 ng de los ARNs purificados, 1 O U/¡JL de la enzima
fosfatasa antártica (New England Biolabs, M0289S), 4 IJL del amortiguador 1 OX (Bis-tris
propano HCI, 50 mM; i+MgCI2, 1 mM; ZnCI2, 0.1 mM; pH 6) y el volumen necesario de
55

CAPITULO 111
agua libre de ARNasas para alcanzar 30 ~L. La reacción se incubó por 60 min a 37 oc, y
se terminó inactivando la enzima a 65 oc por 5 min y finalmente almacenada a -20 oc.
3.2.3 Preparación de membranas para el Northern Blot Reveerso (NBR)
En una membrana de Nylon-N+ (GE, RPN119B) de 12 cm x 9 cm se colocaron 30 pmoles
por sonda de miARNs por cuadricula, dejándola secar a 22 ·e por 40 min . Para efectuar la
inmovilización de las sondas, las membranas fueron colocados en cuatro papeles filtros
Whatman 3 mm, humedecidas con la solución entrecruzante de acuerdo a lo descrito por
Pall y Hamilton (2008) . Se prepararon 24 mL de la solución entrecruzante por membrana
mezclando agua didestilada estéril, 500 mg de 1-etil-3-(3-dimetilaminopropil) carbodiimida
(DEPC) y 245 ~L de metilimidazol12.5 M (pH 8.0). El proceso de inmovilización se realizó
agregando 12 mL de la solución entrecruzante, e iincubando 30 mina 60 oc en un horno
de hibridación. Posteriormente se añadieron 12 mL adicionales de esta solución ,
incubando 1 h a 60 oc. Finalmente, se realizaron dos lavados con agua didestilada por 60
seg cada uno, dejando secar la membrana a 20 ·e por 40 min y almacenándola a -20 ·c.
3.2.4 Análisis por NBR: marcaje radiactivo e hibridación
Una vez que las membranas fueron fijadas e inmovilizadas con la solución entrecruzante,
se procedió a la realización de la prehibridación adicionando 15 mL del amortiguador
comercial UL TRAhyb® (AM8669, Ambion) e incubándolas a 37 oc por 1.5 h. En este
tiempo se realizó la reacción para el marcaje radiactivo, mezclando 20 ~L de agua estéril,
8 ~L del amortiguador 10X T4 Poli-nucleótido Kinasa (PNK), 10 U/~L T4 PNK, 10 ~L de
ARN defosforilado y 1.5 ~L 32P-y-ATP, >7000 Ci/ mmole (-150 ~Ci/~L), incubando a 37
oc por 1 h, seguido de la in activación de la enzima T 4 PNK a 65 oc por 5 m in y 5 m in en
hielo. Una vez que se realizó el proceso de prehibridación y marcaje en la membrana, se
agregaron las muestras marcadas radiactivamente y 5 mL de amortiguador de
prehibridación UL TRAhyb® para iniciar la hibridación a 42 oc por 30 h. Después de
realizar el proceso de hibridación se desechó el amortiguador de hibridación y se
realizaron dos lavados, uno de 3 y otro de 5 min a 42 oc con 15 mL de la solución 5X
SSC/SDS [citrato de sodio/ NaCI (SSC)/0.1% dodecil-sulfato de sodio (SOS)].
Posteriormente, cada membrana fue cubierta con una película plástica (Kieen pack) y se
colocaron en un casete de exposición con dos Hyperfilm ECL (28-9068-35 GE) del
56

CAPITULO 111
tamaño de la membrana en un cuarto oscuro. La exposición se llevó a cabo por 15 días a
-80 oc, para su posterior revelado.
3.2.5 Preparación de las muestras para el análisis del Northern Blot Convencional (NBC)
El análisis mediante NBC es ligeramente diferente al descrito para el NBR. En el caso del
NBR, después de la corrida electroforética, los ARNs pequeños son cortados del gel y
purificados. Por el otro lado, para el análisis del NBC, los ARNs pequeños son
transferidos del gel a una membrana Hybond-N+.
3.2.6 Preparación de las membranas para el NBC
El proceso de electrotransferencia de los ARNs pequeños a las membranas, se realizó
primeramente equilibrando las membranas, las almohadillas Whatman de 0.25 mm y el
gel de la corrida electroforética en el amortiguador de transferencia TBE 1X por 30 min.
Dicho proceso se realizó mediante un ensamblaje en el siguiente orden, apilando las
almohadillas Whatman de 0.25 mm, la membrana Hybond-N+, el gel y almohadillas
Whatman de 0.25 mm en el equipo Simuir transblot (Biorad). Una vez que se retiró el
exceso de amortiguador de transferencia, se selló el apilamiento con una placa metálica y
se realizó la electrotransferencia por 1 h a 1 O V constantes. Una vez terminada la
electrotransferencia la membrana se dejó secar por 40 min a 22 oc. Una vez transcurrido
este tiempo, las membranas fueron fijadas con 12 mL de la solución entrecruzante como
se describió en el protocolo para NBR.
3.2.7 Marcaje radiactivo y proceso de hibridación
Las condiciones para el análisis de las membranas para el NBC son similares a la
realizada para el NBR, lo cual involucra la prehibridación , el marcaje radiactivo y la
hibridación. Las membranas fueron colocadas en los recipientes para realizar la
prehibridación adicionando 1 O mL del amortiguador UL TRAhyb® e incubando durante 1.5
h a 37 oc. Durante este tiempo se efectúo la reacción de marcaje radiactivo mezclando 12
!JL de agua bidestilada estéril , 4 !JL de amortiguador para la enzima 1 OX T4 PNK, 1 !JL de
la enzima T4 PNK, 1 !JL de la sonda del miARN (20 mM) y 1 !JL de la marca 32P-y-ATP,
>7000 Ci/mmole (-100 ¡.JCi/!JL). La mezcla se incubó a 37 oc por 1 h, 5 mina 85 oc y 5
57

CAPITULO 111
min en hielo. A continuación se realizó la prehibridación a 42 oc agregando el volumen
total de la alícuota marcada radiactivamente a la membrana en agitación por 30 h a 60 oc. Los lavados de las membranas se realizaron como se describe para las membranas del
NBR.
3.2.8 Revelado de las películas del NBR y NBC
El revelado de las películas fotográficas se realizó en el cuarto oscuro, retirando las
películas del casete y exponiéndolas a una solución reveladora (Kodak-Sigma-Aidrich
revelador 50/50% agua [v/v]) durante 5 min. A continuación las películas se lavaron con
abundante agua y se transfirieron a la solución fijadora (Kodak-Sigma-Aidrich fijador
aclarador 50/50% agua [v/v]) por 5 min. Seguidamente, las películas se lavaron con
abundante agua y se dejaron secar por 40 min a temperatura ambiente. Posteriormente,
la película fue digitalizada con el equipo Gel Doc XR (Bio-Rad) usando una placa
intensificadora para su posterior análisis digital.
3.2.9. Análisis de la expresión relativa de los miARNs durante la ES
3.2.9.1 Síntesis del ADN complementario (ADNc) y reacción de
poliadenilación
A partir del ARN extraído (tema 2.2.3), los ARNs pequeños fueron purificados mediante
columnas NucleoSpin® miRNA. Las concentraciones fueron verificadas con un Nanodrop
(Thermo Fisher Scientific) y analizadas en geles desnaturalizantes de poliacrilamida al
12.5%. Posteriormente se realizó la síntesis del ADNc a partir de los miARNs utilizando el
kit Ncode® miRNA first strand cONA synthesis (lnvitrogen) , siguiendo las instrucciones del
fabricante. La poliadenilación del ARN purificado se realizó mezclando 2.5 ~g del ARNs, 5
~L de amortiguador de reacción 5X miARNs, 2.5 1..1L de MnCI2 (25mM), 1 ~L de dATP 10
mM, 1 ~L de Poly A polimerasa y agua HyPure grado biología molecular hasta un
volumen final de 25 ~L. La reacción se incubó a 37 oc por 15 m in. Para la síntesis del
ADNc se mezclaron 4 ~L del ARN poliadenilado, 1 ~L del amortiguador de alineamiento y
3 ~L de la secuencia universal RT miRNAs (25 mM); la mezcla de reacción se incubó por
5 m in a 65 oc. A continuación se adicionaron 1 O ¡.~L de 2X first-strand reaction mix y 2 ~L
de la enzima Super-Script 111 RT/ARNsa out, y se incubó por 1 h a 50 oc; la reacción se
58

CAPITULO 111
terminó incubando la mezcla a 85 oc por 5 min y las muestras se almacenaron a -20 oc.
3.2.9.2 Análisis de la expresión de los miARNs por PCR tiempo real (qPCR)
El análisis de expresión por qPCR se realizó utilizando el equipo qPCR cycler (Applied
Biosystems) y el kit Ncode<TM) qRT-PCR (lnvitrogen). Se siguieron las instrucciones
descritas por el fabricante, el ADNc sintetizado fue diluido 1:1 O con agua DEPC. Las
reacciones fueron preparadas a un volumen final de 20 IJL, mezclando 1 O IJL de syber
green qPCR mix, 0.4 IJL de los miARNs de interés (1 O mM), 0.4 IJL del sebador universal
qPCR miRNAs, 0.4 IJL de Passive reference (ROX) y 2 IJL del ADNc (1 :1 O) y 7.2 IJL de
agua tratada con DEPC. Las cond iciones de amplificación para los miARNs fueron 50 oc durante 2 m in, 95 oc durante 1 O m in y 40 ciclos a 95 oc durante 15 seg y 60 oc durante
60 seg.
3.2.9.3 Determinación de los niveles de expresión de los miARNs
El análisis para la determinación de los niveles de expresión relativa de los miARNs se
realizó estableciendo primero la eficiencia de cada miARN y de las curvas de calibración
con las secuencias snU6 y la región 5.8'S. Para determinar la cantidad adecuada de cada
miARN que se usará en el RT-qPCR, se prepararon reacciones con diferentes
concentraciones de ADNc 0.01, 0.1 , 1, 1 O y 1 00 ng en cada una de las muestras (Hj , Mp,
Glo, Cor, Tor, Cot y Cig) y se usaron como testigos de carga al 5.8'S y el snU6. La
determinación de la eficiencia se realizó utilizando la media de los datos obtenidos como
Ct (ciclo umbral) de las reacciones en la curva estándar del testigo endógeno 5.8'S y de
cada miARNs, sustituyendo dichos valores en la ecuación E = 10/\ [-1/valor de la
pendiente]. Posteriormente, para determinar las unidades relativas de expresión se aplicó
la sustitución de datos obtenidos en la ecuación R = [(EmiRNA) ñCt.miRNA] 1 [(E 5.8s) -
ñCt5.8s], en la que ~CT = Ct calibrador - Ct de la muestra ES (en la que para el Ct del
calibrador se utilizó el Ct de snU6 en las muestras de café y 5.8'S se utilizó como testigo
endógeno normalizador).
3.2.1 O Análisis estadístico
Los análisis estadísticos se llevaron a cabo con el software Origin 8.0. Los datos para
determinar la cantidad necesaria para el RT-qPCR de cada miARN fueron obtenidos de la
59

CAPITULO 111
media de dos replicas experimentares y el análisis de expresión relativa de las muestras
problemas fue realizado por triplicado. Los valores fueron evaluados utilizando un análisis
de varianza (ANOVA), donde el grado de significancia entre el valor de las medias se
realizó con la prueba de Tukey con un 95% de confianza.
3.3 RESULTADOS
3.3.1 Análisis del patrón de expresión de los miARNs candidatos por
Northern blot reverso (NBR)
Para responder a la pregunta biológica ¿Cuál es la abundancia de los miARN en las
diferentes etapas del desarrollo embriogénico en C. canephora?, primeramente se realizó
una revisión minuciosa y exhaustiva en las bases datos y las publicaciones hasta el año
2012. Se seleccionaron 47 miARNs con características tanto conservadas como
divergentes en distintas especies vegetales, incluyendo en primera instancia los once
miARNs reportados como conservados en el proceso de la ES.
Los miARNs candidatos durante este trabajo fueron analizados a través de tres técnicas
moleculares, en el primer acercamiento se determinaron patrones de expresión y/o
represión en las muestras del proceso embriogénico de C. canephora, Hj, Mp, Glo, Cor,
Tor, Cot y Cig mediante la técnica del Northern blot reverso (NBR), a partir de estos
resultados se realizó una nueva selección de miARNs, y se determinó cuantitativamente
el patrón de expresión mediante análisis de qPCR-tiempo real. Finalmente, por medio de
la técnica del Northern blot convencional (NBC) se analizaron los patrones particulares de
expresión genética de cinco miARNs, seleccionados entre los miARNs evaluados por
qPCR-tiempo real y NBR.
El patrón de expresiónde los miARNs analizados por NBR se ilustra en el cuadro 3.1 y
muestra los patrones diferenciales de expresión obtenidos para cada miARNs durante las
dos etapas que comprende el proceso de desarrollo embriogénico en C. canephora. En
primer lugar cabe mencionar que por lo menos siete de los miARNs analizados en este
proceso (miR808, miR847t/c, miR1862, miR3623b, miR3629, miR3636 y miR5158) no
fueron detectados mediante esta técnica. Eso puede deberse a que no están presentes o
60

CAPITULO 111
que su expresión está por debajo de los límites de detección de la técnica utilizada. En
segundo lugar se determinó la presencia de una serie de miARNs de una forma
dependiente del tejido y del estadio de desarrollo. En la muestra del explante foliar, día
cero de la inducción de la ES, se detectaron los siguientes miARNs: miR154, miR156,
miR158, miR164, miR165, miR169, miR170, miR172, miR309, miR319, miR390, miR395,
miR397, miR430, miR449, miR535, miR778, miR824, miR827c, miR866 y miR2199.
Después de 21dpi, que corresponde al aislamiento de la Mp, se identificaron a los
miARNs: miR156, miR157, miR160, miR164, miR165, miR170, miR398, miR427, miR449,
miR535, miR778, miR822, miR824, miR2119 y miR3623a. Durante el inicio de la
diferenciación celular y comienzo del desarrollo de las estructuras embriogénicas (estadio
globular) se identificaron los miARNs: miR158, miR163, miR164, miR168, miR170,
miR172, miR395, miR396, miR430, miR535, miR824, miR1524, miR2119, miR2199 y
miR3623a.
Durante la etapa de desarrollo y transición de las estructuras embriogénicas, la dinámica
de expresión de los miARNs expresados fue la siguiente: durante el estadio corazón los
miARNs presentes fueron : miR154, miR156, miR157, miR158, miR159, miR160, miR163,
miR164, miR166, miR167, miR168, miR169, miR170, miR171, miR390, miR393, miR396,
miR397, miR398, miR399, miR449, miR472, miR482, miR535, miR822, miR824, miR827t,
miR866, miR1524, miR2119, miR3623a. Para la transición al estadio torpedo se
expresaron los miARNs: miR154, miR156, miR157, miR158, miR159, miR160, miR164,
miR166, miR167, miR168, miR169, miR170, miR171 , miR309, miR319, miR390, miR393,
miR396, miR395, miR397, miR398, miR399, miR472, miR482, miR535, miR827t, miR866,
miR1524, miR2119, miR3623a. Para el estadio cotiledonar se detectó la expresión de los
miARNs: miR156, miR157, miR158, miR159, miR160, miR164, miR166, miR167, miR168,
miR170, miR171 , miR172, miR309, miR319, miR390, miR393, miR397, miR398, miR396,
miR472, miR482, miR535, miR778, miR822, miR827t, miR1524, miR2119. Finalmente, en
los embriones cigóticos se detectó la expresión de los miARNs: miR154, miR156, miR157,
miR158, miR164, miR165, miR166, miR167, miR169, miR390, miR398, miR399, miR427,
miR472, miR535, miR778, miR2119, y miR3623a.
La evaluación de las dos etapas del proceso embriogénico en C. canephora permitió
determinar la dinámica de la expresión de los miARNs. En el explante foliar usado para la
61

CAPITULO 111
inducción de la ES se expresan 21 miARNs; 21 dpi se detectaron 16 miARNs y al inicio de
la diferenciación celular y establecimiento de los primeros embriones globulares se
detectó la expresión de 15 miARNs. En los estadios Cor, Tor y Cot, se determinó la
expresión de entre 27 y 30 miARNs. También se pudo determinar que hay diferencias
entre los embriones cotiledonares cigóticos y somatícos, pues en los primeros se detectó
la expresión de 18 miARNs, en lugar de los 30 detectados en los somáticos. Estos datos
sugieren la presencia de una dinámica en la expresión de los miARNs durante el proceso
de embriogénesis y permite resaltar las diferencias en los niveles de expresión entre los
dos procesos, el somático y el cigótico.
Los miARNs identificados durante su expresión en la ES fueron agrupados en tres clases,
respecto a los criterios considerados en la etapa de selección, en el primer grupo miARNs
conservados evolutivamente y con funciones regulatorias en la ES de las especies M.
truncatula , L. leptolepis, D. Logan, O. Sativa y V. orange, en tanto que en la segunda
clase se agruparon a los miARNs reportados en los últimos años como miARNs que
participan en la regulación del estrés in vitro, sistemas de micropropagación vegetal y
maduración del embrión cigótico. En la tercera clase se agruparon a los miARNs que
regulan diferentes extremos ambientales en an imales y que por primera vez fueron
analizados durante el proceso de la ES en esta investigación (Cuadro 3.2)
62

CAPITULO 111
Cuadro 3.1 Resultados de expresión cualitaivos por Northern blot reverso. Presencia (+) ausencia (-) de expresión en los miARNs evaluados durante el proceso embriogénico en C. canephora. miARNs conservados (azul) , miARNs implicados en la maduración del embrión cotiledonar y en el estrés in vitro (verde), miARNs candidatos para el proceso de ES (naranja). Masa pro-embriogénica (Mp), embrión globular (Gio) , embrión corazón (Cor), embrión torpedo (Tor) , embrión cotiledonar (Cot) y embrión cigótico (Cig).
miARN Hoja Mp Glo Cor Tor Cot Cig m1 R156 + + - + + + + miR159 - - - + + + -miR160 1 - + - + + + -miR164 1 + + + + + + + miR165 + + - - - - + miR166 - - - + + + + miR167 1 - - - + + + + miR168 1 - - + + + + -miR169 + - - + + - + miR171 - - - + + + -miR172 1 + - + - - + -miR319 1 + - - - + + -miR390 1 + - - + + + + miR393 - - - + + + -miR397 + - - + + + -miR398 1 - + - + + + + miR824 + + + + - - -miR157 - + - + + + + miR396 1 - - + + + + -miR309
1 + - - - + + -
miR427 1 - + - - - - + miR430 1 + - + - - - -miR482 1 - - - + + + -miR535 1 + + + + + + + miR154 - + + + + + -miR158 1 + - + + + + + miR163 - - + + - - -miR170 + + + + + + -miR395 1 + - + - + - -miR399 1 - - - + + - + miR449 1 + + - + - - -miR472 1 - - - + + + + miR778 1 + + - - - + + miR822 1 - + - + - + -miR827t 1 + - - + + + -miR866 1 + - - + + - -miR1524 1 + - - + + - + miR2119 1 - + + + + + + miR2199 + - + - - - -
miR3623a - + + + + - +
63

CAPITULO 111
Cuadro 3.2. Clasificación general de los miARNs presentes durante el proceso embriogénico en C. canephora y analizados por NBR.
Clasificación de los miARNs en C. canephora Hoja Mp Glo Cor Tor Cot
Conservados evolutivamente y reguladores de la ES 8 5 3 13 14 14
Regulan el estrés in vitro y maduración embriogénica 4 4 4 5 5 5
Modulan la respuesta a diferentes extremos ambientales y no han sido reportados en la 9 7 8 13 11 8 e m briogénesis
Total 21 16 15 31 30 27
3.3.2 Determinación de los patrones de expresión por RT-qPCR de 16
miARNs en tejidos embriogénicos de C. canephora
Cig
8
3
7
18
Del análisis de los reportes realizados hasta el momento y del acercamiento experimental
descrito en esta investigación, es clara la participación de los miARNs durante la
inducción y el desarrollo de la ES. Además, con el fin de profundizar y ampliar este
conocimiento durante las fases de inducción y la formación del embrión cotiledonar, se
realizó un análisis más detallado de la presencia de los miARN para determinar su
panorama global durante estos procesos, con base en la primera aproximación obtenida
por NBR.
Con base en los patrones de expresión de los miARN durante estos procesos biológicos,
se realizó la selección de 16 miARNs para la determinación de los patrones de expresión
por RT-qPCR. El grupo de miARNs analizados durante esta etapa incluyó inicialmente a
miR156, miR159, miR164, miR166, miR167, miR168, miR171 , miR390, miR393, miR397
y miR398 que se han reportado como miARN conservados y ejerciendo funciones tejido
específico en la ES de V. orange, también intervienen en la regulación de la maduración
de los embriones cigóticos. Subsecuentemente, se agregaron tres miARNs a esta lista,
los miR157, miR482 y miR535, ya que miR157 se expresa desde Mp, hasta el Cot;
miR482 se expresa en los estadios Cor, Tor y Cot, en tanto que miR535 se expresa en
todo el proceso embriogénico, así como en el embrión cigótico de C. canephora. Es
interesante mencionar que la expresión de estos miARNs también ha sido reportada en
64

CAPITULO 111
procesos de micropropagación masiva y estrés in vitre . Finalmente los miR1524 y
miR2119, analizados previamente por NBR, también fueron considerados ya que se
expresan en la mayor parte del proceso embriogénico y este es el primer trabajo en el
cual, a estos miARN se les ha evaluado como reguladores de la expresión genética
durante la ES, pues particularmente solo se les ha relacionado con el desarrollo radicular.
Para la determinación por RT-qPCR de los 16 miARNs seleccionados, en el orden
anteriormente descrito, se realizó la calibración descrita en materiales y métodos
empleando como testigos internos la expresión de los genes 5.8'S y snU6. También se
determinó la eficiencia del ensayo, mediante el análisis sobre rango dinámico y la
precisión del ensayo realizado para cada miARNs (Anexo 1 ). Los valores obtenidos para
la eficiencia fueron del 86 y 98%, respectivamente.
El análisis de los patrones de expresión de los miARNs conservados: miR156, miR159,
miR164, miR166, miR167, miR168, miR171 , miR390, miR397 y miR398, durante la ES de
C. canephora mostró cierto comportamiento tejido-específico. Durante el análisis en la
etapa inicial del proceso embriogénico se evaluó la expresión de los miARN en explante
foliar, considerado como el día O de la inducción embriogénica. Claramente hay una
diferencia en la expresión de los miR164, miR390 y miR397 (Figuras 3.1 y 3.2). La
expresión más abundante en la de miR390 (2.5 URE) y las expresiones más bajas para
miR164 (0.5 URE) y miR397 (0.2 URE). En la Mp, aislada del explante en el día 28 del
proceso embriogénico, sólo se encuentra presente m iR 164 (1 .0 URE); este miAR N se
expresa más abundantemente en la Mp que en el tejido foliar. Durante la fase de embrión
globular miR164 aumentó (1.75 URE), en tanto que miR168 (0.6 URE) se expresa en
niveles muy bajos (Figuras 3.1, 3.2).
Durante el estadio de Cor se determinó que hay una abundante expresión de miR397 (2.5
URE), miR390 (2.35 URE), miR166 (2.0 URE), miR171 (2 .0 URE). La expresión de
miR168 (1.4 URE), miR393 (1.6 URE) fue intermedia, y para los siguientes miARNs fue
mínima: miR164 (1.0 URE), miR159 (1.0 URE), miR167 (0.9 URE) y miR156 (0.2 URE).
Para el estadio Tor se observó la expresión abundante de miR390 (2.75 URE), miR398
(2 .25 URE), miR159 (2.0 URE) y miR171 (2.0 URE); expresiones intermedias para
miR393 (1.7 URE), miR166 (1 .5 URE), miR167 (1 .0 URE) y miR168 (1 .0 URE) y una
expresión mínima para miR397 (0.4 URE), miR156 (0.5 URE) y miR164 (0.75 URE). Para
65

CAPITULO 111
la transición Tor-Cot se identificaron expresiones abundantes de miR171 (2. 75 URE) y
miR390 (2.5 URE), expresiones intermedias para miR393 (1.8 URE), miR398 (1 .75 URE),
miR167 (1.4 URE) y miR166 (1.25 URE) y expresiones menores para miR164 (1 .0 URE),
miR159 (1.0 URE), miR168 (0.9 URE), miR156 (0.4 URE) y miR397 (0.4 URE). En el caso
del embrión cigótico se determinó que la expresión fue abundante para miR390 (2.35
URE) y miR164 (2.0 URE), intermedia para miR167 (1.1 URE) y menor para miR156 (0.2
URE) (Figuras 3.1 y 3.2) .
El análisis de los patrones de expresión de los miARNs miR157, miR482, miR535,
miR1524 y miR2119 durante el desarrollo de la ES, mostró niveles marcadamente
diferenciados (Figura 3.3) . Durante el explante correspondiente al día cero, se observaron
expresiones abundantes en miR535 (2.20 URE) y miR1524 (2.35 URE}, en tanto que en
el día 28 dpi correspondiente a la Mp, se observarán expresiones intermedias en miR157
(1.18URE) y miR2119 (1 .30 URE}, y minima para miR535 (0.9 URE). Para el desarrollo
del primer estadio embriogénico (Gio) , miR535 (2.20 URE) fue abundante respecto a
miR2119 (1.0 URE) con una expresión intermedia, necesarios para la diferenciación de
este estadio. No obstante, para la transición de los estadios embriogénicos Cor, Tor y Cot
la dinámica entre estos miARNs mostrarán que durante el estadio Cor, miR1524 (2.1
URE) se expresa en mayor abundancia respecto a miR535 (0.25 URE), miR482 (0.8
URE), miR157 (1.0 URE) y miR2119 (1 .9 URE). Durante la transición para el estado Tor,
miR1524 mantuvo su expresión , en tanto que miR482 (2.0 URE}, miR157 (1.20 URE) y
miR535 (0.5 URE) aumentaron su abundancia, en tanto que para miR2119 (1 .1 URE)
disminuyo. Finalmente durante el estadio Cot, la mayor abundancia se presento en
miR157 (2.0 URE), miR535 fue constante y miR482 (0.75 URE), miR1524 (1.75 URE) y
miR2119 (0.3 URE) disminuyendo significativamente. Sin embargo, durante el estadio
cotiledonar cigótico (Cig) se observó la expresión de miR1524 (2.25 URE), miR535 (2.0
URE}, miR157 (1.25 URE) y miR2119 (1 .2 URE).
De acuerdo con los patrones de expresión obtenidos en los siete tejidos analizados en el
proceso embriogénico de C. canephora , los miARNs analizados fueron agrupados
finalmente en tres clases: los miARNs que intervinieron en la primera etapa de inducción
embriogénica (Hj , Mp, Glo) (miR164, miR168, miR390, miR397, miR157, miR535, miR154
y miR2119), los miARNs que ejercieron cierta regulación durante el proceso de transición
66

CAPITULO 111
embriogénica (Cor, Tor, Cot) (miR156, miR159, miR164, miR166, miR167, miR168,
miR171 , miR390, miR397, miR398, miR397, miR157, miR482, miR535, miR1524 y
miR2119) y finalmente los que encontraron durante el estadio Cig (miR164, miR390,
miR156, miR167, miR157, miR535, miR154 y miR2119} , presentando expresiones
minimas, intermedias, abundantes o ausentes en el curso temporal de la ES en C.
canephora (Cuadro 3.3) .
Cuadro 3.3 Patrones de expresión cuantitativa (qPCR-tiempo real) minimas (+), intermedias(++), abundantes (+++) y ausentes (ND) durante el proceso embriogénico de Coffea canephora.
miARNs Hoja Masa pro-
Globular Corazón Torpedo Cotiledonar Cigótico embrigénica
miR156 ND ND ND + + + +
miR159 ND ND ND + +++ ++ ND
miR164 + + ;H + + ++ +++
miR166 ND ND ND +++ ++ ++ ND
miR167 ND ND ND + ++ ++ ++
miR168 ND ND + ++ ++ + ND
miR171 ND ND ND +++ +++ +++ ND
miR390 +++ ND ND +++ +++ +++ +++
miR393 ND ND ND ++ ++ ++ ND
miR397 + ND ND +++ + + ND
miR398 ND ND ND ++ +++ ++ ND
miR157 ND ++ ND ++ ++ +++ ++
miR482 ND ND ND + +++ + ND
miR535 +++ + +++ + + + +++
miR1524 +++ ND ND +++ +++ ++ +++
miR2119 ND ++ ++ ++ ++ + ++ Valores cuant1tat1vos de la Unidades Relat1vas de Expresión (URE): (+) 0.01 - 0.99 URE, (++) 1.0 - 1.99 URE, (+++) 2.0 - 3.0 URE
67

CAPITULO 111
miR156 ..
o.urrollo embtlog6nle0 <10 c . canepltOr•
a.s miR164
•
"' ::0
Desarrollo mb11Q06nleo ele c . canephora
miR167 2,0 ,.. ••• ·~ t.2
&! 1,0
::011,8
1).<1
~
u o
Hj" Hj Mp Glo COt TOf COl Clg
o. .. I'I'OIIo •mbl1ogjnlco <10 C. c•IH>phora
2 ,S
2 ,0
1.1
... "' ::0
1 .0
o,s
o.o
1,0
1,$
"' "' ::0
0,5
O, O
1,11
1,-• . ... 1,2
1,0 111
"' 0,1 ::0
O,t
o 0,2
o,o
m iR159
•
Hj' Hj Mp Olo Cor Tor Cot Clg
OHarroJio embrlogtnleo de c. canaphor•
miR166 *
miR168
l,.,
DH•rrollo embrlogtnlco <10 C. can phora
Figura 3.1. Niveles relativos de expresión de los miARNs conservados en el sistema embriogénico de C. canephora. La expresión relativa (URE) se calculó con base en la .relación [(E miRNA) ACt. miRNA] 1 [(E5.8S) ACt.5.8S]. Para la cual se usaron los valores obtenidos en el análisis de eficiencia de PCR (E), ciclo de la PCR en el que se detecta el producto (Ct) y el gen de referencia (5.8'S). El nivel de expresión snU6 en hoja (Hj*) fue utilizado como normalizador para los miR156, miR159, miR164, miR166, miR167 y miR168. Los asteriscos indican la diferencia significativa de cada miARNs respecto a las muestras analizadas, mediante el análisis estadístico de ANOVA y prueba de Tukey >0.05 error.
68

CAPITULO 111
miR171 miR39 0
miR 393 m R3 97
m iR398
Figura 3.2. Niveles relativos de expresión de miARNs conservados en el sistema embriogénico de C. canephora. La expresión relativa (URE) se tomaron con base en la relación = [(E miRNA) .óCt miRNA] 1 [(E5.8S) .óCt.5.8S]. Para la cual se necesitaron los valores obtenidos en el análisis de eficiencia de PCR (E), ciclo de la PCR en el que se detecta el producto (Ct) y el gen de referencia (5.8'S). El nivel de expresión de snU6 en hoja (Hj*) fue utilizado como normalizador para los miR171 , miR390, miR393, miR397 y miR398. Los asteriscos indican la diferencia significativa de cada miARNs respecto a las muestras analizadas, mediante el análisis estadístico de ANOVA y prueba de Tukey >0.05 error.
69

CAPITULO 111
2.5
U.l 1.5 a:: '::)
o.s
miR157
.Hj" Hj Mp Glo Cor Tor Cot Clg DHarrollo embriogénico de C. canephora
miR535
Hj• Hj: Mp Gto Cor Tor Cot Cig Desarrollo ombrlog6nlco do C. canophora
2 ,1)
1·1·
1,1 1,4
~ u ::::) 1,1>
m iR482
·*
Hj• Hj Mp Glo Cor Tor Cot Cig Desarrollo embriogénico de C. canephora
mi R1524
Hj• Hj Mp Glo Cor Tor Cot. Cig Oe&arrollo embrlogénl<:o ele c. eaMphor<~
miR21 19
Hj* Hj: Mp Glo Cor Tor Cot Cig Desarrollo embriogénico de C; canephora
Figura 3.3. Niveles relativos de expresión de miARNs divergentes en el sistema embriogénico de C. canephora. La expresión relativa (URE) se tomaron con base en la ·relación = [(E miRNA) t.Ct .miRNA] 1 [(E5.8S) t.Ct.5.8S]. Para la cual se necesitaron los valores obtenidos en el análisis de eficiencia de PCR (E), ciclo de la PCR en el que se detecta el producto (Ct) y el gen de referencia (5.8'S). El nivel de expresión snU6 en hoja (Hj*) fue utilizado como normalizador para los miARNs relacionados al estrés in vitro y maduración embriogénica miR157, miR482, miR535 y los miARNs reguladores de estreses ambientales ex vitro miR1524 y miR2119. Los asteriscos indican la diferencia significativa de cada miARNs respecto a las muestras analizadas, mediante el análisis estadístico de ANOVA y prueba de Tukey >0.05 error.
70

CAPITULO 111
3.3.3 Análisis de patrones de expresión semi-cuantitativos por Northern Blot
Convencional (NBC)
A través de la obtención de los datos experimentales sobre los patrones de expresión
parcial por NBR y las expresiones cuantitativas analizadas por RT-qPCR hemos
identificado el papel de los miARNs durante el proceso embriogénico, sin embargo, para
obtener el análisis completo de los miARNs, fue necesario el análisis semi-cuantitativo
mediante NBC. Es decir, a partir de los patrones de expresión identificados por NBR y el
análisis cuantitativos mediante RT-qPCR en los cuales se utilizaron las mismas sondas,
se realizó la selección individual de los miARNs: miR157, miR164, miR166, miR535 y
miR2119 para la validación experimental mediante la hibridación por Northern blot
convencional, con la secuencias de las sondas antisentido marcadas radiactivamente con
[32P] ATP, proporcionando de esta manera el análisis detallados en por lo menos los cinco
miARNs evaluados por las tres técnicas durante el proceso embriogénico de C.
canephora .
Los resultados obtenidos mediante esta técnica mostraron que los niveles de expresión
de miR157 durante el desarrollo embriogénico de C. canephora son abundantes en los
estadios torpedo (Tor) , cotiledonar (Cot) y en el embrión cigótico (Cig) muestran una baja
expresión en la masa pro-embriogénica (Mp) y el estadio de corazón (Cor). También se
determinó que este miARNs no se expresa en el estadio globular (Gio) y el explante foliar
que representa el día cero de la inducción embriogénica (Hj) . Al mismo tiempo miR164 se
expresó durante todo el proceso de la ES; sin embargo, se expresó preferencialmente en
la Mp y en el estadio de Cor, y presenta una expresión constante en los estadios
embriogénicos Glo, Tor, Cot y en el embrión Cig . El miR166 solo se expresa en los
estadios Cor, Tor y Cot y no lo hace en el resto de los tejidos analizados. Por su parte
miR535 se expresó mayormente en Hj y el estadio Glo, en tanto que se expresó en
niveles muy bajos en los estadios Tor y Cot y mientras que su expresión fue constante en
los estadios Mp, Cor y en el embrión Cig. miR2119 se expresó en la Mp, y su mayor
expresión fue en el estadio de Cor, en tanto que su minina expresión se dio en los
estadios Glo y en el embrión Cig, su expresión fue constante en la Mp y en los estadios
Tor y Cot (Figura 3.4) .
Finalmente mediante la comparación de los patrones de expresión de los miARN
71

CAPITULO 111
analizados semi-cuantitativamente, pudimos corroborar el patrón de expresión obtenido
por RT-qPCR de los miR157, miR164, miR166 , miR535 y miR2119 (Figuras 3.1, 3.2 y
3.3), puntualizando que las expresiones analizadas semi-cuantitativas fueron comparadas
con el testigo de carga del ARN nuclear U6 (snU6).
miARNs Gl.o Cor Tor Cot
miR157
miR164
mi-R166
miR535
miH2119
snU6
Figura 3.4. Detección de los miARNs utilizando el análisis Northern blot convencional durante la ES de C. canephora. El análisis de las muestra se realizó por hibridación con sondas correspondientes a miR157, miR164, miR166, miR535, miR2119 marcadas radiactivamente con fosforo [32P] A TP y una exposición de 20 dí as a -80°C. Los tej idos analizados corresponden a hoja (Hj) , masa pro-embriogén ica (Mp), estadios globular (Gio) , corazón (Cor), torpedo (Tor) y cotiledonar somático (Cot) asf como embrión cigótico cotiledonar (Cig), comparados con control de carga snU6.
3.4 Discusión
Hasta la fecha , se han identificado numerosos precursores y secuencias maduras de
miARNs en diversas especies de plantas (www.miRBase.com) . La comprensión sobre la
funcionalidad de los genes, a través de la regulación de los miARNs, depende hoy en día
de la disponibilidad de estrategias innovadoras y del enfoque biológico de la investigación ,
(Jin et al. , 201 0). Actualmente, se han desarrollado diversas metodologías con el objeto
72

CAPITULO 111
de establecer una vía sensible y específica para la detección en el genoma de todos los
miRNAs presentes en las plantas. Los métodos más utilizados para la identificación y la
validación de los miARNs, implican la clonación directa de secuencias maduras de los
miARNs, RT-qPCR cuantitativo, secuenciación masiva y los métodos basados en la
hibridación de sondas por Northern blot, hibridación in situ y microhileras (Eidem et al.,
2013; Jin et al., 201 O).
En conjunto, estas metodologías han generado evidencias sobre la abundancia de los
miARNs durante procesos bilógicos como el ciclo celular, el desarrollo vegetativo y la
respuesta de la plantas ante diversos extremos ambientales; además, estos métodos han
conducido en gran parte a la identificación de los miARN y el destino funcional que estos
ejercen en el nivel de expresión del ARNm, representando así una valiosa información
sobre las funciones de los miARN reportados hasta el momento (Eidem et al. , 2013;
Zhang et al., 2005). En este trabajo, considerando las ventajas e inconvenientes de las
metodologías para la identificación y validación de miARNs, se emplearon las
metodologías basadas en la hibridación de sondas por Northern blot reverso y Northern
blot convencional, así como el análisis cuantitativo por RT-qPCR, con las cuales se
obtuvo una aproximación sobre los patrones de expresión de los miARNs durante la ES
de C. canephora,
En este estudio se identificarán 40 miARNs en los siete tejidos estudiados de C.
canephora. Los patrones de expresión indicán una importante dinámica durante las dos
etapas que conlleva el proceso de la ES (Cuadro 3.1 ). Los patrones de expresión
observados durante el inicio de la inducción, la diferenciación celular y la transición
embriogénica utilizando NBR permitieron observar los patrones de expresión de miARNs
conservados y divergentes (Rhoades, 2012; Zhang et al., 2006a), analizados por primera
vez durante este proceso embriogénico (Cuadro 3.2). Este primer acercamiento permitió
realizar una analogía respecto a los miARNs que han sido reportados en la ES a partir de
callo embriogénico de V. orange (Xiao-Meng et al., 2011) , L. leptolepis (Zhang et al.,
2010; Zhang et al., 2012), D. logan (Lin y Lai, 2013), así como de los embriones de P.
teada aislados durante la embriogénesis somática directa de esta especie (Oh et al.,
2008).
Entre los miARNs analizados hay ciertos miARNs que se expresaron notablemente
73

CAPITULO 111
durante los estadios que comprende la ES y el embrión Cig, en particular miR164 y
miR535. miR164 ha sido ampliamente estudiado en el desarrollo de brotes, meristemos
apicales, plasticidad celular y senescencia de las hojas (Raman et al., 2008; Rubio
Somoza y Weigel, 2011), en tanto que miR535 ha sido estudiado durante el estrés in vitro,
el desarrollo de cultivos de la manera convencional y la micropropagación (Li et al., 2009;
Mica et al., 2009). Otros miARNs, como miR1524 y miR2119, sólo habían sido reportados
durante el desarrollo del sistema radicular (Lelandais-Briére et al., 2009; Subramanian et
al., 2008). Durante este estudio, estos miARNs se expresaron en las fases de transición
embriogénica, diferenciación celular e inducción de la ES, lo que sugiere que
posiblemente estos miARNs tienen otra función biológica. Otros de los miARNs
ampliamente estudiados en varias especies vegetales han sido miR168 y miR396, al
primero se le ha relacionado en el proceso de maduración de los embriones (Mica et al. ,
2009), y miR396 con la proliferación celular (Rodríguez et al., 201 O). En este trabajo se
demostró que estos miARNs se expresan durante el desarrollo de las estructuras
embriogénicas Glo, Cor, Tor y Cot.
Hasta el momento, la expresión de los miARNs analizados en la ES en L. /eptolepis y V.
orange, ha mostrado la expresión de un grupo de 11 miARNs durante el proceso de la ES
indirecta (Zhang et al., 2012; Xiao-Meng et al. , 2011 ). En C. canephora se determinó la
expresión de miR156, miR159, miR166, miR167, miR171, miR390, miR397 y miR398
durante el desarrollo y transición de las estructuras de Cor, Tor y Cot. La expresión de
miR157, miR160, miR393 y miR482 durante estas mismos estadios de la ES sugiere la
participación de miARNs divergentes, a los que se les ha relacionado recientemente con
estreses in vitro (Nonogaki, 201 O; Si-Ammour et al., 2011; Li et al. , 201 O; Rhoades, 2012) .
La expresión de miR309 se ha relacionado con la programación celular (Bushati et al. ,
2008) y la expresión de miR319 con el desarrollo de semillas, hojas, así como a los
extremos de sequia y salinidad (Schommer et al. , 2008; Ori et al., 2007) .
Sin duda, es un panorama muy interesante y ciertamente muy complejo, sin embargo, con
la cuantificación de los miARNs: miR154, miR156, miR157, miR159, miR164, miR166,
miR167, miR168, miR171, miR390, miR393, miR397, miR398, miR482, miR535, miR1524
y miR2119, se expuso un panorama más detallado de la interacción entre los miARNs que
intervinieron en la regulación de la ES tanto de los conservados, como de los divergentes
74

CAPITULO 111
(Figura 3.1), estos últimos poco relacionados con el proceso de ES (Figuras 3.2 y 3.3) ,
pero esenciales para el desarrollo correcto de la ES en C. canephora.
El análisis cuantitativo de estos miARNs, durante los diferentes estadios del proceso
embriogénico, puso en evidencia la presencia de los patrones de expresión de miARNs
necesarios para el desarrollo y regulación de la respuesta de los explantes sometidos al
proceso de inducción embriogénica. Este análisis mostró que hay una baja expresión de
miR164 y miR397, y expresiones abundantes para miR390, miR535 y miR1524.
Indudablemente cada nivel de expresión desempeña un papel clave en la inducción de la
ES; por ejemplo, la expresión de miR164 en A. thaliana regula las concentraciones de
auxinas en la planta, así como la respuesta a etileno y por lo tanto, probablemente, se
encuentra regulando la respuesta al daño mecánico a través de la regulación
transcripcional de NAC1 y ORE1 los que a su vez regulan el proceso de senescencia
foliar (Kim et al. , 2009). miR397, el cual se expresa en bajos niveles, dirige las primeras
repuestas del explante a través de modulación de Lacease (Meng et al., 2012; Axtell y
Bartel, 2005), en tanto que miR390, que regula post-transcripcionalmente la biogénesis de
los elementos actuando en trans, siARNs TAS3, interrumpiendo y degradando el ARNm
que proviene de los genes que codifican para enzimas de la síntesis de auxinas, está
relacionado con miR164 como posible ca-regulador en la concentración de las auxinas
presentes en el tejido foliar (Zhang et al. , 2006b).
Los eventos involucrados en la diferenciación celular de la Mp a Glo, muestran la
dinámica en los niveles de expresión para regular el desarrollo y la competencia celular, a
través de una baja expresión de miR164 y miR535 y una expresión elevada de miR157 y
miR2119 en el desarrollo de la Mp. Durante la formación de la forma esférica y textura lisa
que caracteriza al estadio Glo, los niveles de expresión de miR164 y miR535 aumentaron,
al mismo tiempo que se reprime la expresión de miR157 y mantiene los niveles de
expresión de miR168 y miR2119 a un mínimo. Esta dinámica de regulación y
desregulación entre los miARNs se puede relacionar con las funciones regulatorias
reportadas para los factores CUC1 mediante una baja expresión de miR164. De esta
manera se regula la plasticidad y la diferenciación celular (Rubio-Somoza y Weigel, 2011).
Por su parte, miR157 que ejerce un papel clave sobre la Mp, ha sido funcionalmente
caracterizado en la vía de señalización para inducir el desarrollo embriogénico, mientras
75

CAPITULO 111
que miR168, el cual interviene en la regulación de los factores de respuesta a auxinas
(ARF's) (Kou et al., 2012; Qiao et al. , 2012) , se expresa en niveles altos en C. canephora.
No cabe duda que la regulación ejercida por estos miARNs marcan un papel fundamental
durante el proceso de la diferenciación celular, sin embargo, nuestros resultados no
coinciden en su totalidad con lo reportado durante la ES de V. orange (Xiao-Meng et al.,
2011), L. lepto/epis (Zhang et al., 2012) y O. sativa (Chen et al., 2011) , en particular la
participación de miR156, miR168 y miR171 en la inducción del callo embriogénico, así
como la expresión de miR166, miR167 y miR398 durante la formación del estadio Glo.
Alagunas de estas diferencias podrían estar determinadas por el tipo de explante y la vía
embriogénica utilizada, pues se ha reportado que el tipo de embriogénesis, las
características fenotípicas y el origen del explante influyen en los patrones de expresión
durante la diferenciación celular y el desarrollo de las estructuras embriogénicas (López
Gómez et al. , 201 O; Nassuth et al. , 1980).
Durante el desarrollo de la segunda etapa de la ES, la expresión cuantitativa de los
miARNs: miR156, miR157, miR159, miR164, miR166, miR167, miR168, miR171 , miR390,
miR393, miR397, miR398, miR482, miR535, m iR 1524 y miR2119 mostraron patrones de
expresión diferentes a partir del desarrollo del embrión Glo y el proceso de transición a los
estadios Cor, Tor y Cot. La regulación ejercida por los miARNs durante esta transición ha
sido estudiada en las especies V. orange, D. logan, L. leptolepis, P. teada y O. sativa, en
los cuales se determinó que existe una regulación diferencial de los miARNs involucrados
en la transición embriogénica del estadio Glo al estadio Cot (Xiao-Meng et al. , 2011 ; Lin y
Lai , 2013; Zhang et al. , 2012; Oh et al. , 2008; Zhou et al. , 2010) . En C. canephora esta
dinámica comprendió principalmente niveles de expresión relativamente abundantes para
los miR390 y miR171 , así como niveles mínimos de expresión para miR156 y miR535.
Este patrón de regulación fue similar al reportado previamente para V. orange y P. teada
en los cuales se observaron una expresión abundante de miR390 y miR171 y bajos
niveles de expresión para m iR 156 durante el proceso de transición y desarrollo del
embrión cotiledonar, permitiendo de esta manera una detallada programación para
degradar dinámicamente el ARNm que codifican a los receptores tipo cinanas (RLK) y
TAS3 mediante la acción de miR390, SCL a través de miR171 y SBPy SPL regulados por
m iR 156 (Xiao-Meng et al. , 2011 ; Oh et al., 2008).
76

CAPITULO 111
Un segundo tipo de relación, es la que muestran los miARN, miR166, miR393, miR398 y
m iR 1524, los que se expresan mayormente durante el desarrollo de los primordios
cotiledonares que caracteriza al estadio Cor y una expresión baja durante la transición y el
desarrollo del estadio Cot. Estos resultados son similares a los reportados para miR166,
miR393 y miR398 en D. Logan y L. Jeptolepis, y conforme se van desarrollando los
embriones estos miARNs ejercen una amplia regulación sobre los factores
transcripcionales y genes como APL2 y HD-Zip 111, particularmente por miR166, en tanto
que la expresión de F-box, NF-YC11 y TIR1 es regulada por miR393 y la de la Cu/Zn
superoxide dismutase es regulada por miR398, esto garantiza la transición del estadio Cor
al Cot (Un y Lai , 2013; Zhang et al. , 2012) , Por el contrario, esta relación en la expresión
de los miARNs, durante la transición embriogénica en V. orange es distinta, ya que
durante el desarrollo del estadio globular la expresión de los miARNs se encuentra en
mínimo y alcanzan un máximo durante el desarrollo del embrión cotiledonar (Xiao-Meng et
al. , 2011).
Un tercer tipo de patrón de expresión detectada durante la transición embriogénica fue la
mostrada por los mi ARNs, miR164, miR159 y miR482. La expresión de estos miARNs es
mayor durante el estadio Tor y bajo durnate los estadios de Cor y Cot. Los miANRs,
miR397, miR168 y miR2119 se expresan en mayor abundancia en el estadio de corazón y
la disminuyen durante el desarrollo del embrión Cot, por el contrario los miARNs, miR167
y miR157, se expresan abundantemente en el este estadio. Esta dinámica en la expresión
de los miARNs en C. canephora también se ha determinado en V. orange, P. teada y L.
leptolepis para los miARNs, miR157, miR159, miR164, miR167, miR168 y miR397 (Xiao
Meng et al. , 2011 ; Oh et al. , 2008; Zhang et al. , 2010 ; Zhang et al., 2012).
Los miARNs, miR482, miR535, m iR 1524 y miR2119, analizados por primera vez en este
proceso embriogénico, y que previamente se les ha relacionado con los sistemas de
propagación vegetativo (U et al. , 2009) y el desarrollo del sistema radicular (Subramanian
et al., 2008; Lelandais-Briére et al. , 2009) , se les puede relacionar con la regulación de
algunos de los estadios embriogénicos.
Considerando la dinámica diferencial en la expresión de los miARNs, miR156, miR157,
miR164, miR167, miR390, miR535, miR1524 y miR2119, evaluados cuantitativamente
durante la ES de C. canephora y dada la estrecha semejanza que se presenta entre el
77

CAPITULO 111
desarrollo del embrión somático y la del embrión cigótico, podemos especular que hemos
ampliado nuestra visión sobre la regulación de los miARNs involucrados en la ES y la
regulación del desarrollo del embrión cigótico.
Se ha reportado que en el embrión Cig miR164 regula el desarrollo de los meristemos a
través del control de NACS; también se ha sugerido activa la participación de las auxinas
en el desarrollo vegetativo (Rubio-Somoza y Weigel, 2011 ; Raman et al. , 2008). Por su
parte, miR390 y miR167 son reguladores del desarrollo cotiledonar cigótico, y esta función
se ha relacionado con funciones regulatorias en la división celular mediante la modulación
de los receptores tipo cinasas (RLK) y AUX/AIA (Marin et al. , 2010; Sunkar et al. , 2005).
Por otra parte, m iR 156 y m iR 157 son reguladores de la cascada de señalización en el
desarrollo embriogénico, como lo son SPL y la Squamosa Promoter binding Protein
(SBP), los cuales están implicados en la transición embriogénica y en el desarrollo de
hojas. Los miARNs, miR535, miR1524 y miR2119, que son reportados por primera en
esta investigación se expresan diferencialmente en la mayor parte del proceso
embriogénico estudiado. Finalmente, el estudio de los miARNs, miR157, miR164, miR166,
miR535 y miR2119 en el proceso de ES (Figura 3.4), mostró que participan en la
regulación del proceso embriogénico.
En suma, se ha generado una base sólida para la comprensión funcional y el papel que
juegan en la regulación estos miARNs durante el proceso embriogénico de C. canephora.
Pues aunque esta técnica es ampliamente utilizada para la caracterización de los
miARNs, mayormente se ha utilizado para comprobar funciones específicas de miARN
relacionados con extremos de salinidad y sequia (Zhao et al., 2009; Tran, 2009; Pall y
Hamilton, 2008). En esta investigación se probó que la expresión ·de miR166 en los
estadios Cot y Cig de P. teada (Oh et al. , 2008), muestra exactamente el mismo patrón
que en C. canephora. También podemos concluir que las diferencias en la expresión
temporal de algunos miARNs, como miR164 y miR535, juegan un papel importante en la
inducción de la ES, m iR 157 y miR2119 son esenciales para el proceso de
desdiferenciación y diferenciación celular, en tanto que la expresión de miR166 es
necesaria para el desarrollo temprano y transición entre los estadios de Cor, Tor y Cot.
78

CAPITULO 111
3.5 REFERENCIAS
Adai , A , Johnson, C., Mlotshwa, S., Archer-Evans, S. , Manocha, V., Vanee, V. , y Sundaresan, V. (2005). Computational prediction of miRNAs in Arabidopsis thaliana. Genome Research 15, 78-91 .
Allen, E. , Xie, Z. , Gustafson, A.M., y Carrington, J.C. (2005). microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 121 , 207-221.
Aquea, F. y Arce-Johnson , P. (2008). ldentification of genes expressed during early somatic embryogenesis in Pinus radiata. Plant Physiology and Biochemistry 46, 559-568.
Axtell , M.J., y Bartel , D. P. (2005). Antiquity of microRNAs and their targets in land plants. The Plant Cell 17, 1658-1673.
Bonnet, E. , Wuyts, J., Rouzé, P., y Van de Peer, Y. (2004). Detection of 91 potential conserved plant microRNAs in Arabidopsis thaliana and Oryza sativa identifies important target genes. Proceedings of the National Academy of Sciences of the United States of Ame rica 101 , 11511-11516.
Bushati , N. , Stark, A. , Brennecke, J., y Cohen, S.M. (2008). Temporal reciprocity of miRNAs and their targets during the maternal-to-zygotic transition in Drosophila. Current Biology 18, 501-506.
Chen , C.J., Liu, Q. , Zhang , Y.C. , Qu, L.H ., Chen , Y.Q., y Gautheret, D. (2011). Genomewide discovery and analysis of microRNAs and other small RNAs from rice embryogenic callus. RNA Biology 8, 538-547.
Chen, X. (2005). microRNA biogenesis and function in plants. FEBS Letters 579, 5923-5931.
Cuperus, J.T. , Fahlgren, N. , y Carrington, J.C. (2011). Evolution and functional diversification of miRNA genes. The Plant Cell23, 431-442.
Eldem, V. , Okay, S., y ünver, T. (2013). Plant microRNAs: new players in functional genomics. Turkish Journal of Agriculture and Forestry 37, 1-21.
Eyles, R.P., Williams, P.H., Ohms, S.J., Weiller, G.F., Ogilvie, H.A. , Djordjevic, M.A. , y lmin, N. (2013). microRNA profiling of root tissues and root forming explant cultures in Medicago truncatula. Planta 238, 91-105.
Palatnik, J.F. , Allen, E. , Xuelin , W., Schommer, C. , Schwab, R. , Carrington, J., y Weigel , D. (2003). Control of leaf morphogenesis by microRNAs. Nature 425, 257-263.
Jenik, P.D., Gillmor, C.S., y Lukowitz, W. (2007) . Embryonic patterning in Arabidopsis thaliana. Annual Review of Cell and Developmental Biology 23, 207-236.
Jin , H., Tuo, W., Lian , H., Liu, Q. , Zhu, X.Q. , y Gao, H. (2010). Strategies to identify
79

CAPITULO 111
microRNA targets: New advances. New Biotechnology 27, 734-738.
Karami , 0. , Aghavaisi, B. , y Pour, A.M. (2009). Molecular aspects of somatic-toembryogenic transition in plants. Journal Chemical Biology 2, 177-190.
Khraiwesh, B., Zhu , J.K. , y Zhu , J. (2012). Role of miRNAs and siRNAs in biotic and abiotic stress responses of plants. Biochimica et Biophysica Acta 1819, 137-148.
Kim, J.H ., Woo, H.R. , Kim, J., Lim, P.O., Lee, I.C. , Choi , S.H. , Hwang, D. , y Nam, H.G. (2009). Trifurcate feed-forward regulation of age-dependent cell death involving miR164 in Arabidopsis. Science 323, 1053-1057.
Kou , S.J., Wu, X.M. , Liu , Z., Liu, Y.L., Xu, Q., y Guo, W.W. (2012). Selection and validation of suitable reference genes for miRNA expression normalization by quantitative RT-PCR in citrus somatic embryogenic and adult tissues. Plant Cell Reports 31 , 2151-2163.
Lelandais-Briére, C., Naya, L., Sallet, E., Calenge, F., Frugier, F., Hartmann, C., Gouzy, J., y Crespi , M. (2009). Genome-Wide Medicago truncatula small RNA analysis revealed novel microRNAs and isoforms differentially regulated in roots and nodules. The Plant Cell21 , 2780-2796.
Li , H., Deng , Y. , Wu, T. , Subramanian , S., y Yu , O. (2010). Misexpression of miR482, miR1512, and miR1515 increases soybean nodulation . Plant Physiology 153, 1759-1770.
Li , H., Zhang , Z., Huang, F., Chang , L. , y Ma, Y. (2009). MicroRNA expression profiles in conventional and micropropagated strawberry (Fragaria x ananassa Duch.) plants. Plant Cell Reports 28, 891-902.
Lin, Y. y Lai , Z. (2013). Comparative analysis reveals dynamic changes in miRNAs and their targets and expression during somatic embryogenesis in Longan (Dimocarpus longan Lour). Plos One 8, e60337.
Llave, C. , Xie, Z. , Kasschau, K.O. , y Carrington, J.C. (2002) . Cleavage of Scarecrow-like mRNA targets directed by a class of Arabidopsis miRNA. Science 297, 2053-2056.
López-Gómez, P. , lracheta-Donjuan, L. , Castellanos-Juárez, M., Sandovai-Esquivez, A , Aguirre-Medina, F., Ojeda-Zacarías, M.C., y Gutiérrez-Díez, F. (201 0) . Influencia del explante y medio de cultivo en la embriogénesis somática en hojas de café. Revista Fitotecnia Mexicana 33, 205-213.
Mallanna, S.K. , y Rizzino, A. (201 O). Emerging roles of microRNAs in the control of embryonic stem cells and the generation of induced pluripotent stem cells. Developmental Biology 344, 16-25.
Mallory, A.C. , y Vaucheret, H. (2006). Functions of microRNAs and related small RNAs in plants. Nature Genetics 38, S31-S36.
80

CAPITULO 111
Marin, E. , Jouannet, V., Herz, A. , Lokerse, A.S. , Weijers, D., Vaucheret, H., Nussaume, L. , Crespi, M.O. , y Maizel, A. (2010) . miR390, Arabidopsis TAS3 tasiRNAs, and their AUXIN RESPONSE FACTOR targets define an autoregulatory network quantitatively regulating lateral root growth. The Plant Cell 22, 1104-1117.
Meng, Y., Shao, C. , Ma, X., Wang , H., y Chen, M. (2012). Expression-based functional investigation of the organ-specific microRNAs in Arabidopsis. Plos One 7, e50870.
Mica, E., Piccolo, V. , Delledonne, M., Ferrarini , A. , Pezzotti, M. , Casati, C. , Del Fabbro, C., Valle, G. , Policriti, A. , Morgante, M., Pesole, G., Pe, M.E., y Horner, D. (2009). High throughput approaches reveal splicing of primary microRNA transcripts and tissue specific expression of mature microRNAs in Vitis vinifera. BMC Genomics 10, 558.
Nassuth, A. , Wormer, T.M., Bouman, F., y Staritsky, G. (1980). The histogenesis of callus in Coffea canephora stem explants and the discovery of early embryoid initiation. Acta Botanica Neerlandica 29, 49-54.
Nellikunnuma, S.M. y Chandrashekar, A. (2012). Computational identification of conserved miRNA and their targets in Coffea canephora by EST Analysis. Dynamic Biochemistry Process Biotechnology and Molecular Biology 70.
Nonogaki, H. (201 0). MicroRNA gene regulation cascades during early stages of plant development. Plant and Cell Physiology 51 , 1840-1846.
Oh, T.J. , Wartell , R.M. , Cairney, J. , y Pullman, G.S. (2008). Evidence for stage-specific modulation of specific microRNAs (miRNAs) and miRNA processing components in zygotic embryo and female gametophyte of loblolly pine (Pinus taeda). New Phytologist 179, 67-80.
Ori , N. , Cohen, A.R., Etzioni , A., Brand, A. , Yanai , 0 ., Shleizer, S., Menda, N., Amsellem, Z., Efroni , 1. , Pekker, 1. , Alvarez, J.P. , Blum, E. , Zamir, D., y Eshed, Y. (2007). Regulation of LANCEOLA TE by miR319 is required for compound-leaf development in tomato. Nature Genetics 39, 787-791.
Pall, G.S. y Hamilton, A.J. (2008). lmproved northern blot method for enhanced detection of small RNA. Nature Protocols 3, 1077-1084.
Pei , H., Ma, N., Chen, J., Zheng, Y., Tian, J., Li , J., Zhang , S., Fei , Z., y Gao, J. (2013). lntegrative analysis of miRNA and mRNA profiles in response to ethylene in rose petals during flower opening . Plos One 8, e64290.
Qiao, M., Zhao, Z., Song, Y., Liu, Z., Cao, L. , Yu , Y. , Li, S., y Xiang , F. (2012) . Proper regeneration from in vitro cultured Arabidopsis thaliana requ ires the microRNAdirected action of an auxin response factor. The Plant Journal 71, 14-22.
Raman , S., Greb, T. , Peaucelle, A. , Blein, T., Laufs, P., y Theres, K. (2008). lnterplay of miR164, CUP-SHAPED COTYLEDON genes and LATERAL SUPPRESSOR controls axillary meristem formation in Arabidopsis thaliana . The Plant Journal 55,
81

CAPITULO 111
65-76.
Rhoades, M.W., Reinhart, B.J., Lim, L.P., Burge, C.B., Bartel , B. , y Bartel , O.P. (2002). Prediction of plant microRNA targets. Cell 110, 513-520.
Rhoades, M.W. (2012) . Conservation and divergence in plant microRNAs. Plant Molecular Biology 80, 3-16.
Rodríguez, R.E. , Mecchia, M.A. , Debernardi , J.M., Schommer, C., Weigel, D., y Palatnik, J.F. (2010). Control of cell proliferation in Arabidopsis tha/iana by microRNA miR396. Development 137, 103-112.
Rogers, K., y Chen, X. (2013). Biogenesis, turnover, and mode of action of plant microRNAs. The Plant Cell 25, 2383-2399.
Rubio-Somoza, 1., y Weigel, D. (2011) . MicroRNA networks and developmental plasticity in plants. Trends in Plant Science 16, 258-264.
Schommer, C., Palatnik, J.F., Aggarwal , P., Chetelat, A. , Cubas, P. , Farmer, E.E., Nath, U., y Weigel, D. (2008). Control of jasmonate biosynthesis and senescence by miR319 targets. Plos Biology 6, e230.
Si-Ammour, A. , Windels, D., Bouldoires, E., Kutter, C., Ailhas, J., Meins, F., y Vazquez, F. (2011 ). miR393 and secondary siR NAs regulate expression of the TIR1/AFB2 auxin receptor clade and auxin-related development of Arabidopsis leaves. Plant Physiology 157, 683-691 .
Subramanian , S., Fu, Y., Sunkar, R. , Barbazuk, W.B., Zhu, J.K., y Yu, O. (2008). Novel and nodulation-regulated microRNAs in soybean roots. BMC Genomics 9, 160.
Sunkar, R., Girke, T. , Jain, P.K., y Zhu , J.K. (2005). Cloning and characterization of microRNAs from rice. The Plant Cell 17, 1397-1411.
Thakur, V. , Wanchana, S., Xu, M., Bruskiewich , R. , Quick, W.P., Mosig , A. , y Zhu , X.G. (2011). Characterization of statistical features for plant microRNA prediction. BMC Genomics 12, 108.
Tran, N. (2009) . Fast and simple micro-RNA northern blots. Biochemistry lnsights 2, 1-3.
Tretter, E.M., Alvarez, J.P., Eshed, Y., y Bowman, J.L. (2008). Activity range of Arabidopsis small RNAs derived from different biogenesis pathways. Plant Physiology 147, 58-62.
Wang , J.W., Wang , L.J., Mao, Y.B., Caí, W.J. , Xue, H.W., y Chen , X.Y. (2005). Control of root cap formation by MicroRNA-targeted auxin response factors in Arabidopsis. The Plant Cell 17, 2204-2216.
Wang , W.X. , Gaffney, B., Hunt, A.G., y Tang , G. (2001). MicroRNAs (miRNAs) and plant development. In eLS, John Wiley and Sons, Ltd .
82

CAPITULO 111
Wang, X.J., Reyes, J.L. , Chua, N.H., y Gaasterland, T. (2004). Prediction and identification of Arabidopsis thaliana microRNAs and their mRNA targets. Genome Biology 5, R65.
Willmann, M.R. , Mehalick, A.J. , Packer, R. L., y Jenik, P.D. (2011 ). MicroRNAs regulate the timing of embryo maturation in Arabidopsis. Plant Physiology 155, 1871-1884.
Xiao-Meng , Wu. , Mei-Ya, L. , Xiao-Xia, Ge., Qiang , Xu ., y Wen-Wu, G. (2011). Stage and tissue-specific modulation of ten conserved miRNAs and their targets during somatic embryogenesis of Valencia sweet orange. Planta 233, 495-505.
Xu , L., y Huang, H. (2014). Genetic and epigenetic controls of plant regeneration. Current Topics in Developmental Biology 108, 1-33.
Yang , X. , y Zhang , X.L. (2010). Regulation of somatic embryogenesis in higher plants. Critica! Reviews in Plant Sciences 29, 36-57.
Zhang, B., Pan, X., Cannon , C.H. , Cobb, G.P. , y Anderson, T.A. (2006a). Conservation and divergence of plant microRNA genes. The Plant Journal 46, 243-259.
Zhang, B. , Pan, X. , Cobb, G.P. , y Anderson, T.A. (2006b). Plant microRNA: A small regulatory molecule with big impact. Developmental Biology 289, 3-16.
Zhang , B.H., Pan, X.P. , Wang , Q.L., Cobb, G.P. , y Anderson, T.A. (2005). ldentification and characterization of new plant microRNAs using EST analysis. Cell Research. 336-360.
Zhang, J. , Zhang, S. , Han, S., Li , X., Tong, Z., y Qi , L. (2013). Deciphering small noncoding RNAs during the transition from dormant embryo to germinated embryo in Larches (Larix leptolepis) . Plos One 8, e81452.
Zhang, J., Zhang , S., Han, S., Wu, T., Li , X. , Li , W., y Qi , L. (2012). Genome-wide identification of microRNAs in larch and stage-specific modulation of 11 conserved microRNAs and their targets during somatic embryogenesis. Planta 236, 647-657.
Zhang, S., Zhou, J. , Han, S., Yang, W., Li, W., Wei, H., Li, X., y Qi , L. (2010). Four abiotic stress-induced miRNA families differentially regulated in the embryogenic and nonembryogenic callus tissues of Larix leptolepis. Biochemical and Biophysical Research Communications 398, 355-360.
Zhao, B., Ge, L. , Liang , R. , Li , W., Ruan, K., Lin , H., y Jin, Y. (2009) . Members of miR-169 family are induced by high salinity and transiently inhibit the NF-Y A transcription factor. BMC Molecular Biology 10, 29.
Zhou, L., Liu, Y., Liu , Z., Kong , D., Duan, M., y Luo, L. (2010) . Genome-wide identification and analysis of drought-responsive microRNAs in Oryza sativa. Journal Experimental Botany 61, 4157-4168.
83


CAPITULO IV
CAPITULO IV: DISCUSIÓN GENERAL, CONCLUSIONES Y PERSPECTIVAS
4.1 DISCUSIÓN GENERAL
Las plantas son organismos sésiles que presentan continuo crecimiento en todo su ciclo
de vida, son capaces de generar respuestas rápidas para adaptarse ante diversos
extremos ambientales mediante la plasticidad celular, única de las células vegetales
(Zimmerman, 1993; Yang y Zhang , 201 O). Bajo condiciones controladas el cultivo in vitro
es una herramienta biotecnológica utilizada para la regeneración de plantas, la
conservación del germoplasma y la obtención de plantas genéticamente mejoradas en
diferentes especies agronómicamente importantes (Sachs, 1991 ; Yeung, 1995). Entre las
técnicas in vitro más utilizadas para estos fines está la organogénesis y la embriogénesis,
en donde la plasticidad celular y la capacidad totipotente durante el proceso embriogénico
han sido estudiadas desde los aspectos bioquímicos, moleculares y genéticos. Sin
embargo, en la última década la regulación mediante los mecanismos epigenéticos y
específicamente la regulación ejercida a través de los pequeños ARNs, ha empezado a
ganar terreno para comprender su importancia en los procesos biológicos, por ejemplo, el
desarrollo de la embriogénesis somática (ES).
Los estudios realizados con miARNs durante la embriogénesis han demostrado el papel
clave que estas pequeñas moléculas desempeñan durante este proceso. Por ejemplo,
actualmente se han identificado miARNs en diferentes especies vegetales involucrados en
la inducción embriogénica, la morfogénesis y la maduración del embrión cigótico y
somático (Xiao-Meng et al., 2011; Aquea y Arce-Johnson , 2008; Eyles et al. , 2013) . En la
presente tesis fue posible evaluar 11 miARNs conservados y 29 miARNs divergentes
(Cuadro 3.1.) , los cuales fueron considerados a partir de los miARNs reportados en las
especies S. lycopersicum (Zhang et al. , 2008; Pilcher et al., 2007), G. max (Li et al. , 2011 ;
Chen et al., 2009) , P. trichocarpa (Barakat et al. , 2007; Chen et al., 2011 ; Lu et al., 2005),
V. vinífera (Lu et al. , 2008; Tillett et al. , 2011), M. truncatula (Zhou et al., 2008; lmin et al. ,
2008), V. orange (Xiao-Meng et al., 2011), A. thaliana (Tretter et al. , 2008; Todesco et al.,
85

CAPITULO IV
201 0), así como C. canephora y C. arabica (Nellikunnuma y Chandrashekar 2012).
En general , los resultados obtenidos de la expresión cuantitativa de 11 miARNs
conservados (Figura 3.1, 3.2) y cinco miARNs seleccionados (Figura 3.3), de los 29
miARNs divergentes relacionados por primera vez al proceso de inducción, el desarrollo y
la maduración embriogénica, mostraron ciertos patrones de expresión que sugieren su
participación en el proceso embriogénico de C. canephora. Por ejemplo, la expresión
abundante de miR164, encontrada en el embrión globular y su abundancia mínima
durante todo el proceso de la ES (Figura 3.1 ), podría relacionarse con la regulación de los
genes NAC durante el desarrollo del embrión globular y la maduración del embrión
cigótico como ha sido reportado en A. thaliana (Kim, et al. , 2009; Raman et al. , 2008). Por
lo cual se podría inferir que este miARN se encuentra relacionado al proceso del
desarrollo globular cigótico y somático, así también, podría considerarse como un miARNs
esencial durante el proceso embriogénico en C. canephora.
Otros miARNs conservados y evaluados en este estudio fueron los miARNs; miR156,
miR166, miR167, miR168, miR171 y miR398, reportados hasta el momento como
miARNs involucrados en la ES indirecta y en la regulación de los genes SCARECROW
LIKE-3 (SCL), SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) y
ARGONAUTA-1 (AG01) , importantes durante la inducción y el mantenimiento del callo
embriogénico (Lin y Lai , 2013; Zhang et al. , 2012 ; Xiao-Meng et al. , 2011) . El análisis
encontrado en la primera etapa en la ES (explante foliar y Mp) en C. canephora (Cuadro
3.1) mostró que estos miARNs estuvieron ausentes, lo cual sugiere que los genes SCL,
SPL y AG01 podrían estar expresados en estos tejidos. Además, se ha reportado que
estos genes están involucrados en la diferenciación celular para la generación de la masa
pro-embriogénica (Lin y Lai , 2013). Por otro lado, los resultados obtenidos de miR168
(Figura 3.1) durante la transición embriogénica Glo, Cor, Tor, y Cot de C. canephora,
podría estar relacionada a la regulación de AG01 y RNA-INDUCED TRANSCRIPTIONAL
SILENCING COMPLEX PROTEIN-3 (TAS3) como ha sido reportado en D. logan y L.
leptolepis en donde este miARN ejerce una regulación durante el desarrollo y la transición
de las estructuras embriogénicas deCora Cot (Vaucheret et al. , 2006; Zhang et al. , 2010;
Zhang et al. , 2012, Lin y Lai , 2013).
Algunos de los miARN nuevos analizados en C. canephora como los miR157, miR390 ,
86

CAPITULO IV
miR535, m iR 1524 y miR2119 evaluados durante el proceso embriogénico presentaron
expresiones diferenciales durante la ES (Cuadro 3.3). Por ejemplo, el miR157 y el miR390
se encontraron con expresiones abundantes en los estadios Cor, Tor y Coten la ES de C.
canephora (Cuadro 3.3) . Hasta el momento el miR157 ha sido reportado como un
elemento clave para el desarrollo morfogénico y la transición de fases vegetativas
(Gandikota et al., 2007; Rhoades et al., 2002), mientras que el miR390 ha sido
considerado un regulador clave para el desarrollo cotiledonar, cigótico y el proceso de la
división celular a través de la regulación de los receptores tipo cinasas (RLK) y
AUXIN/INDOLE-3-ACETIC AGIO (AUX/AIA) (Marin et al. , 2010; Sunkar et al. , 2005).
Estos dos miARNs han sido recientemente relacionados con miR535, como posibles
reguladores de los extremos in vitro en especies micropropagados como Strawberry spp
(Li et al. , 2009). Ante estas evidencias y con base a los resultados obtenidos en la ES de
C. canephora (Cuadro 3.3) , se podría sugerir que estos miARNs se encuentran ejerciendo
funciones similares y tal vez podrían estar estableciendo un sistema de autorregulación in
vitro en los factores transcripcionales y genes involucrados en la ES, que dirigen el
desarrollo y transición de los embriones somáticos (Figuras 3.2 y 3.3).
La información generada en este trabajo y la aplicación de las técnicas moleculares
utilizadas han permitido realizar un primer acercamiento sólido para la comprensión
funcional de 16 miARNs durante el proceso embriogénico de C. canephora (Cuadro 3.3) .
Los resultados generados pueden ser considerados el punto de partida de otras
investigaciones que busquen esclarecer el funcionamiento biológico de genes blancos
regulados por estos miARNs y dar respuesta a algunas de las interrogantes sobre el
proceso embriogénico somático y cigótico, así como propuestas biotecnológicas que den
soluciones a algunas de las problemáticas que implican algunos sistemas in vitro
vegetales como la recalcitrancia. Con los resultados obtenidos en este trabajo y la
información disponible en la literatura se puede proponer finalmente un modelo
epigenético sobre el posible mecanismo de acción de los miARNs durante el proceso de
la ES de C. canephora (Figura 4.1 ).
87

CAPITULO IV
AG01
NAC
miR172 ~ APL2
miR4Q~AUX/AII MYB APL ~. ~? ~
familias miR159/miR169/miR171
A miR156 miR1 71
...... miR166
miR2119?
miR535 ?
miR154?
miR168 r AG09 ......-- m::::::::=- B miR16~ HO-ZIPIII
miR15~ MYB33
miR156~ SPL
? siARNs
,-------i ARF17 miR160/miR167 IAUXINASJ
4~&13
miR160/mÍR390 4 ARF2/ARF3/ARF4
Familia miR396
r--1 Represión
-:;;::- Balance para dos miARNs Abundancia elevada
miR396a 4 CDS
miR397 Y LACCASE ~ miR24 ?
TIR ? miR393 --=----' SCL/SCR
Transcripción .A. Auto-regulación
Balance entre varios miARNs ? Blancos de acción desconocidos
Figura 4.1 . Modulación epigenética de los miARNs en la embriogénesis somática de Coffea canephora. Durante el inicio del proceso embriogénico, las células somáticas presentes en el explante foliar experimentan una reprogramación en su epigenoma a través de una regulación ejercida por miARNs conservados y divergentes. En el presente modelo se sugiere la regulación dinámica de 16 miARNs evaluados durante el explante foliar, la masa pro-embriogénica (Mp) y el desarrollo de las estructuras embriogénicas (Gio, Cor, Tor, Cot). (A) Indica la regulación de los
88

CAPITULO IV
miARNs miR154, miR164, miR168 miR390, miR482 y miR535, evaluados por primera vez, durante el proceso de la ES y la posible regulación sobre HEME ACTIVATOR PROTE/N-2 (HAP2), ARGONAUTA1 (AG01) y NAM, ATAF y CUC (NAC). (8) Indica la dinámica ejercida de los miARNs conservados, miR156, miR159, miR160, miR166, miR168, miR171 , miR390, miR393 miR397 y miR398, en la ES de C. canephora sobre el desarrollo y la transición embriogénica a través de la posible regulación de los genes SQUAMOSA PROMOTER 8/ND/NG PROTEIN (SPL), CLASS 111 HOMEODOMAIN-LEUC/NE ZIPPER (HD-Z/P/11), ARGONAUTA (AGO), AUXIN RESPONSE FACTOR (ARF}, SCARECROW-LIKE PROTEIN (SCUSCR}, COPPER SUPEROXIDASE DISMUTASES (CDS), LACCASE, MYB, y TIR como ha sido encontrado en las especies Larix leptolepis, Valencia orange y Dimocarpus /ogan .
4.2 CONCLUSIÓN GENERAL
El principal objetivo de este trabajo fue la identificación de los miARNs presentes durante
el proceso de ES en C. canephora, así como, la determinación de sus patrones de
expresión a lo largo del proceso embriogénico. Los resultados obtenidos sugieren que el
mecanismo epigenético regulado por los miARNs, en el proceso embriogénico de C.
canephora, podría estar altamente regulado por estas pequeñas moléculas desde la
inducción de la ES, hasta el desarrollo y transición entre los diferentes estadios
embriogénicos. Las conclusiones generales de este trabajo son:
~ Se determinó que la transición de las células somáticas, presentes en el explante
foliar, a células con capacidad embriogénica es dirigida por los miRNAs: miR156,
miR164, miR165, miR169, miR172, miR309, miR319, miR390, miR397, miR430,
miR535 y miR824.
~ Se determinó que los miARNs: miR156, miR157, miR160, miR164, miR165,
miR171 , miR398, miR427, miR535 y miR824 regulan el desarrollo de la Mp.
~ Se demostró que para el desarrollo morfogénico y la transición entre los diferentes
estadios embriogénicos es necesaria la participación temprana de los miR164,
miR168, miR172 , miR396, miR430, miR535 y miR824, (estadio globular) , seguida
de la activación de 22 miARNs para la regulación de la transición entre los
estadios corazón , torpedo y cotiledonar. En el embrión cigótico se idéntico la
presencia de niveles bajos de expresión de once miARNs.
~ Se identificaron cuantitativamente los patrones de expresión de 16 miARNs
durante los diferentes estadios del proceso embriogénico de C. canephora. Se
89

CAPITULO IV
determinó la expresión de miR157, miR164, miR168, miR390, miR397, miR535 y
miR1524, durante la inducción y la formación de la masa pro-embriogénica, en
tanto que durante el desarrollo y transición entre los diferentes estadios
embriogénicos se determinó la expresión de miR156, miR157, miR159, miR164,
miR166, miR167, miR168, miR171 , miR390, miR393, miR397, miR398, miR482,
miR535 y miR2119.
>-- La expresión de miR157, miR164, miR166, miR535 y miR2119 se demostró a
través de tres metodologías, lo que permitió establecer los patrones de expresión
de estos miARNs de una forma tejido-especifico.
4.3 PERSPECTIVAS DEL TRABAJO
90
>-- En trabajos futuros será necesario continuar el análisis de los patrones de
expresión relativa durante el proceso de ES en C. canephora con el fin de
identificar nuevos miARNs que puedan estar participando en la regulación del
proceso de ES.
>-- En estudios posteriores será necesario identificar los genes blanco de la acción de
los miARNs durante el proceso de ES en C. canephora .
>-- Será necesario la elaboración de una base de datos para todos los miARNs
relacionados con el proceso de ES.
>-- También será necesario la elucidación de los precursores biológicos de estos
miARNs e identificación de las isoformas que los miARNs pueden presentar.

CAPITULO IV
4.4 REFERENCIAS
Aquea, F., y Arce-Johnson , P. (2008). ldentification of genes expressed during early somatic embryogenesis in Pinus radiata. Plant Physiology and Biochemistry 46, 559-568
Barakat, A. , Wall , P.K. , Diloreto, S., Depamphilis, C.W., y Carlson , J.E. (2007). Conservation and divergence of microRNAs in Populus. BMC Genomics 8, 481-496.
Chen, L. , Zhang, Y. , Ren, Y. , Xu, J. , Zhang , Z. , y Wang, Y. (2011). Genome-wide identification of cold-responsive and new microRNAs in Populus tomentosa by high-throughput sequencing . Biochemical and Biophysical Research Communication 417, 897-896.
Chen, R. , Hu, Z., y Zhang, H. (2009). ldentification of microRNAs in wild soybean (Giycine soja). Journal of lntegrative Plant Biology 51, 1071-1079.
Eyles, R.P., Williams, P.H., Ohms, S.J. , Weiller, G.F. , Ogilvie, H.A. , Djordjevic, M.A. , y lmin,N. (2013). microRNA profiling of root tissues and root forming explant cultures in Medicago truncatula . Planta 238, 91-105.
Gandikota, M., Birkenbihl , R.P. , Hohmann, S., Carden , G.H. , Saedler, H., y Huijser, P. (2007). The miRNA 156/157 recognition element in the 3' UTR of the Arabidopsis SBP box gene SPL3 prevents early flowering by translational inhibition in seedlings. The Plant Journal 49, 683-693.
lmin, N. , Goffard, N., Nizamidin , M., y Rolfe, B.G. (2008). Genome- wide transcriptional analysis of super-embriogenic Medicago truncatula explants cultures. BMC Plant Biology 8-11 O.
Kim, J.H. , Woo,H.R. , Kim,J., Lim,P.O., Lee,I.C., Choi ,S.H ., Hwang ,D., y Nam,H.G. (2009). Trifurcate feed-forward regulation of age-dependent cell death involving miR164 in Arabidopsis. Science 323, 1053-1057.
Li , H. , Dong, Y., Yin , H., Wang, N. , Yang, J. , Liu , X., Wang , Y., Wu, J., y Li , X. (2011). Characterization of the stress associated microRNAs in Glycine max by deep sequencing. BMC Plant Biology 11, 170.
Li , H., Zhang , Z. , Huang, F., Chang, L. , y Ma, Y. (2009). MicroRNA expression profiles in conventional and micropropagated strawberry (Fragaria x ananassa Duch .) plants. Plant Cell Reports 28, 891 -902.
Lin, Y., y Lai, Z. (2013). Comparative analysis reveals dynamic changes in miRNAs and their targets and expression during somatic embryogenesis in Longan (Dimocarpus longan Lour). Plos One 8, e60337.
Lu, S., Sun, Y.H. , Shi , R. , Clark, C., Li, L., y Chiang , V.L. (2005). Novel and mechanical stress-responsive microRNAs in Populus trichocarpa that are absent from
91

CAPITULO IV
Arabidopsis. The Plant Cell17, 2186-2203.
Lu, Y., Gan, Q. , Chi , X. , y Qin, S. (2008). ldentification and characterization of microRNAs and their targets in grapevine (Vitis vinífera). Agricultura! Sciences in China 7, 929-943.
Marin, E. , Jouannet, V., Herz, A., Lokerse, A.S. , Weijers, D. , Vaucheret, H., Nussaume, L. , Crespi , M.O. , y Maizel, A. (2010) . miR390, Arabidopsis TAS3 tasiRNAs, and their AUXIN RESPONSE FACTOR targets define an autoregulatory network quantitatively regulating lateral root growth. The Plant Cell 22, 1104-1117.
Nellikunnuma,S.M. y Chandrashekar,A. (2012). Computational identification of conserved miRNA and their targets in Coffea canephora by EST Analysis. Dynamic Biochemistry, Process Biotechnology and Molecular Biology 6, 70-76.
Pilcher, R.L., Moxon, S. , Pakseresht, N., Moulton , V., Manning, K. , Seymour, G. , y Dalmay, T. (2007). ldentification of novel small RNAs in tomate (Solanum lycopersicum). Planta 226, 709-717.
Raman , S., Greb, T., Peaucelle, A. , Blein , T., Laufs, P., y Theres, K. (2008). lnterplay of miR164, CUP-SHAPED COTYLEDON genes and LATERAL SUPPRESSOR controls axillary meristem formation in Arabidopsis thaliana. The Plant Journal 55, 65-76.
Rhoades, M.W. , Reinhart, B.J., Lim, L.P., Surge, C.B. , Bartel, B., y Bartel , O.P. (2002). Prediction of plant microRNA targets. Cell 110, 513-520.
Sachs, T. (1991). Developmental and cell biology series. Pattern formation in plant tissues. (Cambridge: Cambridge University Press).
Sunkar, R. , Girke, T., Jain, P.K., y Zhu , J.K. (2005). Cloning and characterization of microRNAs from rice. The Plant Cell 17, 1397-1411 .
Tillett, R.L. , Ergul , A. , Albion , R.L. , Schlauch , K.A. , Cramer, G.R. , y Cushman, J.C. (2011). ldentification of tissue-specific, abiotic stress-responsive gene expression patterns in wine grape (Vitis vinífera L.) based on curation and mining of large-scale EST data sets. BMC Plant Biology 11 , 86.
Todesco, M., Rubio-Somoza, 1. , Paz-Ares, J. , y Weigel , D. (201 0). A collection of target mimics for comprehensive analysis of microRNA function in Arabidopsis thaliana . Plos Genetics 6, e1 001031.
Tretter, E.M. , Alvarez, J.P., Eshed, Y., y Bowman, J.L. (2008). Activity range of Arabidopsis small RNAs derived from different biogenesis pathways. Plant Physiology 147, 58-62.
Vaucheret, H. , Mallory, A.C., y Bartel , O.P. (2006) . AG01 homeostasis entails coexpression of MIR168 and AG01 and preferential stabil ization of miR168 by AG01. Molecular Cell 22, 129-136.
92

CAPITULO IV
Xiao-Meng, Wu. , Mei-Ya, L. , Xiao-Xia, Ge. , Qiang , Xu ., y Wen-Wu, G. (2011) . Stage and tissue-specific modulation of ten conserved miRNAs and their targets during somatic embryogenesis of Valencia sweet orange. Planta 233, 495-505.
Yang , X. y Zhang , X.L. (2010). Regulation of somatic embryogenesis inhigher plants. Critica! Reviews in Plant Sciences 29, 36-57.
Yeung , E.C. (1995). Structural and development patters in somatic embryogenesis. In In Vitro Embryogenesis in Plants, Thorpe T.A. Ed: Kluwer Academic Publishers, Netherlans 205-24 7.
Zhang , J. , Zeng , R. , Chen, J. , Liu , X ., y Liao, Q. (2008) . ldentification of conserved microRNAs and their targets from Solanum lycopersicum Min. Gene 423, 1-7.
Zhang , J. , Zhang , S., Han, S., Wu , T. , Li, X. , Li , W., y Qi, L. (2012) . Genome-wide identification of microRNAs in larch and stage-specific modulation of 11 conserved microRNAs and their targets during somatic embryogenesis. Planta 236, 647-657.
Zhang, S. , Zhou, J., Han, S. , Yang , W., Li , W. , Wei , H. , Li, X., y Qi , L. (201 0) . Four abiotic stress-induced miRNA families differentially regulated in the embryogenic and nonembryogenic callus tissues of Larix leptolepis. Biochemical and Biophysical Research Communications 398, 355-360.
Zhou, Z.S. , Huang, S.Q., y Yang, Z.M . (2008) . Bioinformatic identification and expression analysis of new microRNAs from Medicago truncatula . Biochemical and Biophysical Research Communication 374, 538-542.
Zimmerman, J.L. (1993). Somatic embryogenesis: A model for early development in higher plants. The Plant Cell 5, 1411-1423.
93

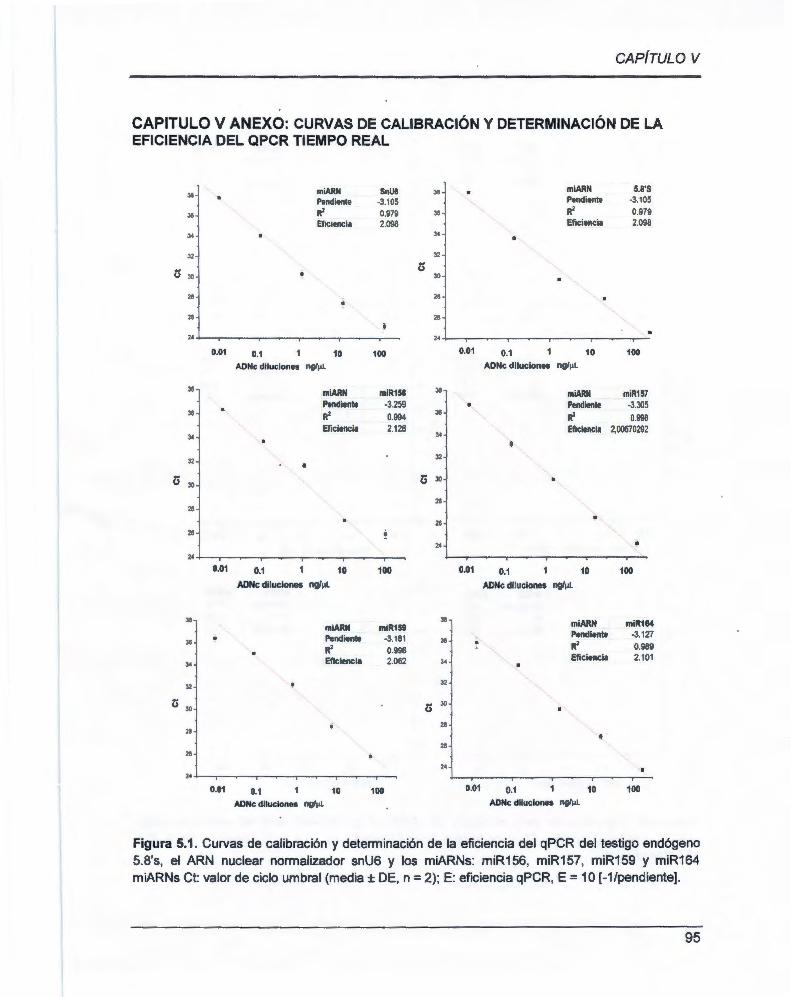
CAPÍTULO V
CAPITULO V ANEXO: CURVAS DE CALIBRACIÓN Y DETERMINACIÓN DE LA EFICIENCIA DEL QPCR TIEMPO REAL
miARN SnU6 38 miAR N 5.8'S 38
Pendiente -3.105 Pendiente -3.1 05
36 R' 0.979 36 R' 0.979 Eficiencia 2.098 Eficiencia 2.098
34 34
32 32
tS 30 tS
30
28 28
28 26
24 24
0.01 0.1 10 100 0.01 0.1 10 100
ADNc diluciones ng/~L AONc diluciones ngii!L
38 miARN miR156 38 miARN miR157 Pendiente -3.259 Pendiente -3.305
36 R' 0.994 38
R' 0.998 Eficiencia 2.126 Eficiencia 2,00670292
34 34
32 32
i3 i3 30 30
2S 2S
26
28
24
24
0.01 0.1 10 100 0.01 0.1 10 100
AONc diluciones ng/¡ll AONc diluciones ng/j!L
38 38 miARN miR164 miARN miR159 Pendiente -3.181 .. Pendiente -3.t27
38 36 . R' R' 0.996 0.989
Eficiencia 2.062 34 Eficiencia 2.101 34
32 32
tS ü 30 30
28 28
28 28
24 24
0.01 0.1 10 100 0.01 0.1 10 100
AONc diluciones ng/1.1L ADNc diluciones ng/~L
Figura 5.1 . Curvas de calibración y determinación de la eficiencia del qPCR del testigo endógeno 5.8's, el ARN nuclear normalizador snU6 y los miARNs: miR156, miR157, miR159 y miR164 miARNs Ct: valor de ciclo umbral (media± DE, n = 2); E: eficiencia qPCR, E= 10 [-1/pendiente].
95

CAPÍTULO V
:l8 38 miARN miR166 miARN miR167
J6 Pendiente -3.157 38 Pendiente -3.305 R' 0.998 R' 0.997
34 Eficiencia 2.073 34 Eficiencia 2.006
32 32
o so ü 30
28 28
26 26
"' 24
0.01 0.1 10 100 0.01 0.1 10 100
ADNc diluciones ng/¡!L AONc diluciones ng/¡.tl
38 miAR N miR168 38 miAR N miR171 Pendiente -3.301 Pendiente -3.030
38 ~ 0.994 38 R' 0.999 Eficiencia 2.009 Eficiencia 2.134
34 34
32 32 ü
30 ! ü 30
28 28
26 26
24 24
0.01 0.1 10 100 0.01 0.1 10 100 AONc diluciones ng/¡.¡L AONc diluciones ng/¡.tl
38 38 miARN miR390 miARN miR393
38 Pendiente -3.035 38 Pendiente -3.067
R' 0.973 R' 0.998
34 Eficiencia 2.321 34 Eficiencia 2.118
32 32
JO ü 30 ü
28 28
26 26
24 24
0.01 0.1 10 100 0.01 0.1 10 100
ADNc diluciones ng/¡.tl AONc diluciones ng/¡.tl
Figura 5.2. Determinación de la eficiencia del qPCR de los miARNs: miR166, miR167 miR168, miR171 , miR390 y miR393. Ct: valor de ciclo umbral (media ± DE, n = 2); E: eficiencia qPCR, E = 10 [-1/pendiente].
96

CAPÍTULO V
"" miARN miR398 miARN miR397 "" Pendiente -3.122 Pendiente -3.098
R' 36 R' 0.994
36 0.997 Eficiencia 2.053 Eficiencia 2.091
34 34
32 32
ü ü
"" JO
28 28
26 26 ..
0.01 0.1 10 100 0.01 0.1 10 100 ADNc diluciones ng/IJ.L ADNc diluciones ng/IJ.L
38 miARN miR482 38
Pendiente -3.051 miARN miR535
R' 0.976 38 Pendiente -3.135
38 R'
Eficiencia 2.126 0.986
34 34 Eficiencia 2.1 84
32 32
ü JO ü JO
28 28
26 26
24
24
0.01 0.1 10 100 0.01 0.1 10 100
ADNc diluciones ng/IJ.L ADNc diluciones ng/IJ.L
38 miARN miR1524
Pendiente 38 miARN miR2119 .-3.235
Pendiente -3.451 38 R2 0.9745 36 R' 0.975
34 Eficiencia 2.084 Eficiencia 1.998 34
32 32
ü )1) ü 30
28 28
26 26
24 24
0.01 0.1 10 100 0.01 0.1 10 100
ADNc diluciones ng/IJ.L ADNc diluciones ng/¡J.L
Figura 5.3. Determinación de la eficiencia del qPCR del los miARNs: miR397, miR398, miR482, miR535, miR1524y miR2119 Ct: valor de ciclo umbral (media ± DE, n = 2); E: eficiencia qPCR, E = 10 [-1/pendiente]
97








